
Fluorescensmikroskop - Fluorescence microscope


Et fluorescensmikroskop er et optisk mikroskop, der bruger fluorescens i stedet for, eller ud over, spredning , refleksion og dæmpning eller absorption for at studere egenskaberne af organiske eller uorganiske stoffer. "Fluorescensmikroskop" refererer til ethvert mikroskop, der bruger fluorescens til at generere et billede, uanset om det er en enkel opsætning som et epifluorescensmikroskop eller et mere kompliceret design, såsom et konfokalt mikroskop , som bruger optisk snitning for at få bedre opløsning af fluorescensbilledet .
Princip
Prøven belyses med lys med en bestemt bølgelængde (eller bølgelængder), som absorberes af fluoroforerne , hvilket får dem til at udsende lys med længere bølgelængder (dvs. af en anden farve end det absorberede lys). Belysningslyset adskilles fra den meget svagere udsendte fluorescens ved brug af et spektralemissionsfilter. Typiske komponenter i et fluorescensmikroskop er en lyskilde ( xenonbue-lampe eller kviksølvdamplampe er almindelig; mere avancerede former er LED'er og lasere med høj effekt ), excitationsfilteret , det dikroiske spejl (eller dikroiske stråleplitter ) og emissionen filter (se figuren nedenfor). Filtrene og den dikroiske strålefordeler vælges for at matche spektral excitations- og emissionsegenskaberne for fluoroforen, der bruges til at mærke prøven. På denne måde afbildes fordelingen af en enkelt fluorofor (farve) ad gangen. Flerfarvebilleder af flere typer fluoroforer skal sammensættes ved at kombinere flere enkeltfarvebilleder.
De fleste fluorescensmikroskoper i brug er epifluorescensmikroskoper, hvor excitation af fluoroforen og påvisning af fluorescensen sker gennem den samme lyssti (dvs. gennem målet). Disse mikroskoper er meget udbredt i biologi og er grundlaget for mere avancerede mikroskopdesign, såsom konfokalmikroskopet og det totale interne refleksionsfluorescensmikroskop (TIRF).
Epifluorescensmikroskopi

Størstedelen af fluorescensmikroskoper, især dem, der bruges inden for biovidenskaben , er af epifluorescensdesignet vist i diagrammet. Lyset af excitationsbølgelængden belyser prøven gennem objektive linse. Den fluorescens, der udsendes af prøven, er fokuseret på detektoren af det samme formål, der bruges til excitationen, som for større opløsning vil have brug for objektiv med en højere numerisk blænde . Da det meste af excitationslyset transmitteres gennem prøven, når kun reflekteret excitatorisk lys målet sammen med det udsendte lys, og epifluorescensmetoden giver derfor et højt signal-til-støj-forhold. Dichroic beamsplitter fungerer som et bølgelængdespecifikt filter, der sender fluoresceret lys igennem til okularet eller detektoren, men reflekterer eventuelt resterende excitationslys tilbage mod kilden.
Lyskilder
Fluorescensmikroskopi kræver intens, næsten monokromatisk belysning, som nogle udbredte lyskilder, som halogenlamper ikke kan levere. Der bruges fire hovedtyper af lyskilder, herunder xenon-buelamper eller kviksølvdamplamper med et excitationsfilter , lasere , superkontinuumkilder og lysdioder med høj effekt . Lasere bruges mest til mere komplekse fluorescensmikroskopiteknikker som konfokalmikroskopi og total intern refleksionsfluorescensmikroskopi, mens xenonlamper og kviksølvlamper og lysdioder med et dikroisk excitationsfilter normalt bruges til videfelt epifluorescensmikroskoper. Ved at placere to mikrolens- arrays i belysningsvejen for et vidfelt-epifluorescensmikroskop kan der opnås en meget ensartet belysning med en variationskoefficient på 1-2%.
Prøveforberedelse
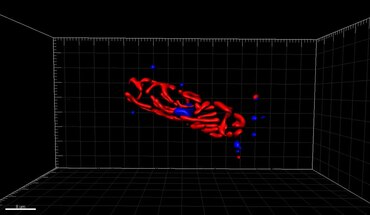
Viser overlejringer fra fire fluorescerende kanaler
kernecellelegemerne (b) Cyan: [PLL -A546 fluorescens] - generisk modfarve til visualisering af eukaryote celleoverflader
(c) Blå: [Hoechst fluorescens] - pletter DNA, identificerer kerner
(d) Rød: [klorofyl autofluorescens] - løser kloroplaster

For at en prøve skal være egnet til fluorescensmikroskopi, skal den være fluorescerende. Der er flere metoder til at oprette en fluorescerende prøve; de vigtigste teknikker er mærkning med fluorescerende pletter eller, i tilfælde af biologiske prøver, ekspression af et fluorescerende protein . Alternativt kan den iboende fluorescens af en prøve (dvs. autofluorescens ) anvendes. Inden for biovidenskaben er fluorescensmikroskopi et kraftfuldt værktøj, der tillader specifik og følsom farvning af en prøve for at detektere fordelingen af proteiner eller andre molekyler af interesse. Som et resultat er der en bred vifte af teknikker til fluorescerende farvning af biologiske prøver.
Biologiske fluorescerende pletter
Mange fluorescerende pletter er designet til en række biologiske molekyler. Nogle af disse er små molekyler, der er iboende fluorescerende og binder et biologisk molekyle af interesse. Store eksempler på disse er nukleinsyrepletter som DAPI og Hoechst (begejstret af UV -bølgelængdelys) og DRAQ5 og DRAQ7 (optimalt exciteret af rødt lys), som alle binder den mindre rille af DNA og dermed mærker cellernes kerner . Andre er lægemidler, toksiner eller peptider, der binder specifikke cellulære strukturer og er blevet derivatiseret med en fluorescerende reporter. Et hovedeksempel på denne klasse af fluorescerende pletter er phalloidin , som bruges til at plette actinfibre i pattedyrceller . Et nyt peptid, kendt som Collagen Hybridizing Peptide , kan også konjugeres med fluoroforer og bruges til at plette denaturerede kollagenfibre. Farvning af plantens cellevægge udføres ved hjælp af pletter eller farvestoffer, der binder cellulose eller pektin . Jakten på fluorescerende sonder med en høj specificitet, der også tillader levende billeddannelse af planteceller, er i gang.
Der er mange fluorescerende molekyler kaldet fluoroforer eller fluorokromer, såsom fluorescein , Alexa Fluors eller DyLight 488 , som kan være kemisk knyttet til et andet molekyle, der binder målet for interesse i prøven.
Immunfluorescens
Immunfluorescens er en teknik, der anvender den meget specifikke binding af et antistof til dets antigen for at mærke specifikke proteiner eller andre molekyler i cellen. En prøve behandles med et primært antistof specifikt for molekylet af interesse. En fluorofor kan konjugeres direkte til det primære antistof. Alternativt kan et sekundært antistof konjugeret til en fluorofor, der specifikt binder til det første antistof, anvendes. For eksempel kan et primært antistof, der er rejst i en mus, som genkender tubulin kombineret med et sekundært antimus-antistof derivatiseret med en fluorofor, bruges til at mærke mikrotubuli i en celle.
Fluorescerende proteiner
Den moderne forståelse af genetik og de tilgængelige teknikker til at ændre DNA gør det muligt for forskere at genetisk modificere proteiner til også at bære en fluorescerende proteinreporter. I biologiske prøver giver dette en forsker mulighed for direkte at lave et protein af interesse fluorescerende. Proteinplaceringen kan derefter spores direkte, herunder i levende celler.
Begrænsninger
Fluoroforer mister deres evne til at fluorescere, da de belyses i en proces kaldet fotoblegning . Fotoblegning sker, når de fluorescerende molekyler akkumulerer kemisk skade fra elektronerne, der ophidses under fluorescens. Fotoblegning kan i høj grad begrænse den tid, over hvilken en prøve kan observeres ved fluorescensmikroskopi. Der findes flere teknikker til at reducere fotoblegning såsom brug af mere robuste fluoroforer, ved at minimere belysning eller ved at bruge fotobeskyttende kemikalier til opfangning .
Fluorescensmikroskopi med fluorescerende reporterproteiner har muliggjort analyse af levende celler ved fluorescensmikroskopi, dog er celler modtagelige for fototoksicitet, især med lys med kort bølgelængde. Desuden har fluorescerende molekyler en tendens til at generere reaktive kemiske arter, når de er under belysning, hvilket forbedrer den fototoksiske effekt.
I modsætning til transmitterede og reflekterede lysmikroskopiteknikker tillader fluorescensmikroskopi kun observation af de specifikke strukturer, der er blevet mærket for fluorescens. For eksempel afslører observation af en vævsprøve fremstillet med en fluorescerende DNA -plet ved fluorescensmikroskopi kun organiseringen af DNA'et i cellerne og afslører intet andet om cellemorfologierne.
Beregningsteknikker, der foreslår at estimere det fluorescerende signal fra ikke-fluorescerende billeder (såsom brightfield) kan reducere disse bekymringer. Generelt involverer disse tilgange uddannelse af et dybt konvolutionsnervalt netværk på farvede celler og derefter estimering af fluorescensen på ufarvede prøver. Ved at frakoble de undersøgte celler fra de celler, der bruges til at træne netværket, kan billeddannelse udføres hurtigere og med reduceret fototoksicitet.
Sub-diffraktionsteknikker
Lysets bølgetype begrænser størrelsen af det sted, som lyset kan fokuseres på grund af diffraktionsgrænsen . Denne begrænsning blev beskrevet i det 19. århundrede af Ernst Abbe og "begrænser et optisk mikroskops opløsning til cirka halvdelen af bølgelængden af det anvendte lys." Fluorescensmikroskopi er centralt for mange teknikker, der har til formål at nå forbi denne grænse ved specialiserede optiske konfigurationer.
Flere forbedringer i mikroskopiteknikker er blevet opfundet i det 20. århundrede og har resulteret i øget opløsning og kontrast til en vis grad. Men de overvandt ikke diffraktionsgrænsen. I 1978 er de første teoretiske ideer blevet udviklet til at bryde denne barriere ved hjælp af et 4Pi mikroskop som et konfokalt laserscannende fluorescensmikroskop, hvor lyset ideelt set fokuseres fra alle sider til et fælles fokus, der bruges til at scanne objektet ved 'punkt-for- punkt 'excitation kombineret med' punkt-for-punkt 'detektion. Den første eksperimentelle demonstration af 4pi-mikroskopet fandt imidlertid sted i 1994. 4Pi-mikroskopi maksimerer mængden af tilgængelige fokusretninger ved at bruge to modstående objektiver eller to-foton-excitationsmikroskopi ved hjælp af rødt forskudt lys og multi-foton-excitation.
Integreret korrelativ mikroskopi kombinerer et fluorescensmikroskop med et elektronmikroskop. Dette gør det muligt at visualisere ultrastruktur og kontekstuel information med elektronmikroskopet, mens data fra fluorescensmikroskopet bruges som et mærkeværktøj.
Den første teknik til virkelig at opnå en sub-diffraktionsopløsning var STED-mikroskopi , foreslået i 1994. Denne metode og alle teknikker, der følger RESOLFT- konceptet, er afhængige af en stærk, ikke-lineær interaktion mellem lys og fluorescerende molekyler. Molekylerne drives stærkt mellem adskilte molekylære tilstande på hvert specifikt sted, så endelig lys kan udsendes på kun en lille brøkdel af rummet, derfor en øget opløsning.
Også i 1990'erne er der blevet udviklet en anden superopløsningsmikroskopimetode baseret på bredfeltsmikroskopi. Betydeligt forbedret størrelsesopløsning af cellulære nanostrukturer farvet med en fluorescerende markør blev opnået ved udvikling af SPDM -lokaliseringsmikroskopi og den strukturerede laserbelysning (rumligt moduleret belysning, SMI). Kombination af princippet om SPDM med SMI resulterede i udviklingen af Vertico SMI -mikroskopet . Enkeltmolekyldetektion af normale blinkende fluorescerende farvestoffer som grønt fluorescerende protein (GFP) kan opnås ved hjælp af en videreudvikling af SPDM den såkaldte SPDMphymod-teknologi, som gør det muligt at detektere og tælle to forskellige fluorescerende molekyltyper på molekylært niveau (dette teknologi kaldes tofarvet lokaliseringsmikroskopi eller 2CLM).
Alternativt kunne fremkomsten af fotoaktiveret lokaliseringsmikroskopi opnå lignende resultater ved at stole på blink eller skift af enkeltmolekyler, hvor fraktionen af fluorescerende molekyler er meget lille hver gang. Denne stokastiske respons af molekyler på det påførte lys svarer også til en meget ulinær interaktion, hvilket fører til subdiffraktionsopløsning.
Fluorescensmikrografgalleri

En z-projektion af en osteosarcomacelle, farvet med phalloidin for at visualisere aktinfilamenter. Billedet blev taget på et konfokalmikroskop, og den efterfølgende dekonvolution blev udført ved hjælp af en eksperimentelt afledt punktspredningsfunktion.
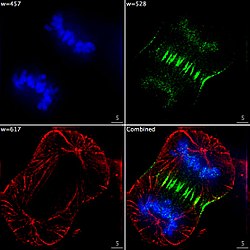
Epifluorescerende billeddannelse af de tre komponenter i en delende human kræftcelle. DNA er farvet blåt, et protein kaldet INCENP er grønt, og mikrotubuli er røde. Hver fluorofor afbildes separat ved hjælp af en anden kombination af excitations- og emissionsfiltre, og billederne fanges i rækkefølge ved hjælp af et digitalt CCD -kamera og derefter overlejres for at give et komplet billede.

Endotelceller under mikroskopet. Nuclei farves blå med DAPI, mikrotubuli er markeret grønt med et antistof bundet til FITC og actinfilamenter er mærket rødt med phalloidin bundet til TRITC. Kvæg pulmonal arterie endotel (BPAE) celler
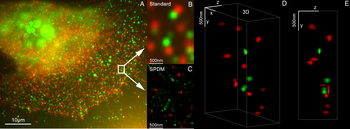
3D dobbeltfarvet superopløselig mikroskopi med Her2 og Her3 i brystceller, standardfarvestoffer: Alexa 488, Alexa 568. LIMON-mikroskopi

Human lymfocytkerne farvet med DAPI med kromosom 13 (grøn) og 21 (rød) centromersonder hybridiseret ( Fluorescerende in situ hybridisering (FISH))

Gærcellemembran visualiseret af nogle membranproteiner smeltet med RFP- og GFP -fluorescerende markører. Påføring af lys fra begge markører resulterer i gul farve.

Superopløselig mikroskopi: Detektion af enkelt YFP-molekyle i en human kræftcelle. Typiske afstandsmålinger i 15 nm-området målt med et Vertico-SMI/SPDMphymod-mikroskop

Superopløselig mikroskopi: Co-lokaliseringsmikroskopi (2CLM) med GFP- og RFP-fusionsproteiner (kerne i en knoglekræftcelle) 120.000 lokaliserede molekyler i et bredfeltområde (470 µm 2 ) målt med et Vertico-SMI/SPDMphymod mikroskop

Fluorescensmikroskopi af DNA-ekspression i Human Wild-Type og P239S Mutant Palladin .

Fluorescensmikroskopibilleder af solblusser -patologi i en blodcelle, der viser de berørte områder med rødt.










Se også
- Fluorescens billeddannelse
- Fluorescens i biovidenskaben
- Korrelativ lys-elektronmikroskopi
- Elizabeth Harry , pioner inden for fluorescensmikroskopiteknikker til visualisering af bakterielle subcellulære proteiner
- Grønt fluorescerende protein (GFP)
- Kviksølvdampe
- Mikroskop
- Scannende elektronmikroskop#katodoluminescens
- Stokes skifter
- Xenon lysbue
Referencer
eksterne links
|
Biblioteksressourcer om fluorescensmikroskopi |
- Fluorophores.org , databasen over fluorescerende farvestoffer
- Mikroskopi Ressource Center
- animationer og forklaringer på forskellige typer mikroskoper, herunder fluorescerende og konfokale mikroskoper (Université Paris Sud)