
Dictyostelium discoideum -Dictyostelium discoideum
| Dictyostelium discoideum | |
|---|---|

|
|
| Frugtlegemer af D. discoideum | |
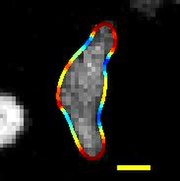
| En migrerende D. discoideum, hvis grænse er farvet med krumning, skala bar: 5 µm, varighed: 22 sekunder | |
|
Videnskabelig klassificering |
|
| Domæne: | Eukaryota |
| Phylum: | Amoebozoa |
| Klasse: | Dictyostelia |
| Bestille: | Dictyosteliida |
| Familie: | Dictyosteliidae |
| Slægt: | Dictyostelium |
| Arter: |
D. discoideum
|
| Binomisk navn | |
|
Dictyostelium discoideum
Raper , 1935
|
|

Dictyostelium discoideum er en art af jordlevende amøber, der tilhører phylum Amoebozoa , infraphylum Mycetozoa . Almindeligvis omtalt som slimform er D. discoideum en eukaryot, der overgår fra en samling encellede amøber til en flercellet snegl og derefter til en frugtlegeme inden for sin levetid. Dens unikke aseksuelle livscyklus består af fire faser: vegetativ, aggregering, migration og kulmination. D. discoideums livscykluser relativt kort, hvilket muliggør rettidig visning af alle faser. Cellerne, der er involveret i livscyklussen, gennemgår bevægelse, kemisk signalering og udvikling, som kan anvendes til forskning i menneskelig kræft. Enkelheden i dets livscyklus gør D. discoideum til en værdifuld modelorganisme til at studere genetiske, cellulære og biokemiske processer i andre organismer.
Naturligt levested og kost
I naturen kan D. discoideum findes i jord og fugtigt bladaffald. Dens primære kost består af bakterier , såsom Escherichia coli , der findes i jorden og henfaldende organisk stof. Uninucleate amøber af D. discoideum forbruge bakterier, der findes i deres naturlige habitat, som omfatter løvskov jord og rådnende blade.
Livscyklus og reproduktion
Den livscyklus af D. discoideum begynder, når sporer frigives fra en moden sorocarp (frugtlegemer organ). Myxamoebae udklækkes fra sporer under varme og fugtige forhold. I løbet af deres vegetative fase deler myxamoebæerne sig med mitose, når de lever af bakterier. Bakterierne udskiller folsyre , som tiltrækker myxamoebae. Når forsyningen af bakterier er opbrugt, går myxamoebæerne ind i aggregeringsfasen.
Under aggregering starter sult produktionen af proteinforbindelser, såsom glycoproteiner og adenylylcyclase . Glykoproteinerne muliggør adhæsion af celler og celler , og adenylylcyklase skaber cyklisk AMP . Cyklisk AMP udskilles af amøberne for at tiltrække naboceller til et centralt sted. Når de bevæger sig mod signalet, støder de ind i hinanden og hænger sammen ved hjælp af glycoprotein -adhæsionsmolekyler.
Migreringsfasen begynder, når amøberne har dannet et tæt aggregat, og den langstrakte cellehøj vælter for at ligge fladt på jorden. Amøberne arbejder sammen som et bevægeligt pseudoplasmodium, også kendt som en snegl. Sneglen er cirka 2–4 mm lang, sammensat af op til 100.000 celler og er i stand til at bevæge sig ved at producere en celluloseskede i dens forreste celler, som sneglen bevæger sig igennem. En del af denne kappe efterlades som et slimet spor, når den bevæger sig mod tiltrækningsmidler som lys , varme og fugtighed i en fremadrettet retning. Cyklisk AMP og et stof kaldet differentieringsinducerende faktor hjælper med at danne forskellige celletyper. Sneglen bliver differentieret til prestalk- og prespore -celler, der bevæger sig til henholdsvis den forreste og bageste ende. Når sneglen har fundet et passende miljø, danner sneglens forreste ende frugtlegemets stilk, og den bageste ende danner frugtlegemets sporer. Forreste-lignende celler, som først for nylig er blevet opdaget, er også dispergeret i hele den bageste region af sneglen. Disse forreste-lignende celler danner selve bunden af frugtlegemet og sporenes hætter. Efter at sneglen har sat sig på ét sted, breder den bageste ende sig ud med den forreste ende hævet i luften og danner det, der kaldes "mexicansk hat", og kulminationstrinnet begynder.
Prestalk -cellerne og presporecellerne skifter positioner i kulminationstrinnet for at danne den modne frugtlegeme. Den forreste ende af den mexicanske hat danner et celluloserør, som gør det muligt for de mere bageste celler at bevæge sig op ad ydersiden af røret til toppen, og fortalcellerne bevæger sig ned. Denne omlægning danner frugtlegemets stilk, der består af cellerne fra sneglens forreste ende, og cellerne fra den bageste ende af sneglen er på toppen og danner nu frugtlegemets sporer. Ved afslutningen af denne 8-10 timers proces er den modne frugtlegeme fuldt ud dannet. Denne frugtlegeme er 1-2 mm høj og kan nu starte hele cyklussen igen ved at frigive de modne sporer, der bliver til myxamoebae.
Seksuel reproduktion
Generelt, selvom D. discoideum generelt gengiver aseksuelt , D. discoideum er stadig i stand til seksuel reproduktion , hvis visse betingelser er opfyldt. D. discoideum har tre forskellige parringstyper, og undersøgelser har identificeret kønlokus, der specificerer disse tre parringstyper. Type I -stammer er specificeret af genet kaldet MatA, Type II -stammer har tre forskellige gener: MatB (homolog til Mat A), Mat C og Mat D, og Type III -stammer har Mat S- og Mat T -gener (som er homologe med Mat C og Mat D). Disse køn kan kun parre sig med de to forskellige køn og ikke med sine egne.
Når de inkuberes med deres bakterielle fødevareforsyning, kan der forekomme heterothallisk eller homothallisk seksuel udvikling, hvilket resulterer i dannelsen af en diploid zygote. Heterothallisk parring opstår, når to amøber af forskellige parringstyper er til stede i et mørkt og vådt miljø, hvor de kan smelte sammen under aggregering for at danne en kæmpe zygotecelle. Kæmpecellen frigiver derefter cAMP for at tiltrække andre celler og opslugter derefter de andre celler kannibalistisk i aggregatet. De forbrugte celler tjener til at indkapsle hele aggregatet i en tyk, cellulosevæg for at beskytte det. Dette er kendt som en makrocyst . Inde i makrocysten deler den gigantiske celle først gennem meiose , derefter gennem mitose for at producere mange haploide amøber, der frigives til fodring, som normale amøber ville. Homothalliske D. discoideum -stammer AC4 og ZA3A er også i stand til at producere makrocyster. Hver af disse stammer udtrykker i modsætning til heterothalliske stammer sandsynligvis begge parringer af alleler ( matA og mata ). Selvom seksuel reproduktion er mulig, er det meget sjældent at se vellykket spiring af en D. discoideum macrocyst under laboratorieforhold. Ikke desto mindre er rekombination udbredt inden for D. discoideum naturlige befolkninger, hvilket indikerer, at sex sandsynligvis er et vigtigt aspekt af deres livscyklus.
Brug som modelorganisme
Fordi mange af dets gener er homologe med menneskelige gener, men dens livscyklus er enkel, bruges D. discoideum almindeligvis som en modelorganisme . Det kan observeres på organismisk, cellulært og molekylært niveau primært på grund af deres begrænsede antal celletyper og adfærd og deres hurtige vækst. Det bruges til at studere celledifferentiering, kemotaksi og apoptose , som alle er normale cellulære processer. Det bruges også til at studere andre aspekter af udviklingen, herunder cellesortering, mønsterdannelse, fagocytose, motilitet og signaltransduktion. Disse processer og udviklingsaspekter er enten fraværende eller for vanskelige at se i andre modelorganismer. D. discoideum er nært beslægtet med højere metazoaner. Det bærer lignende gener og veje, hvilket gør det til en god kandidat til gen -knockout .
Den celledifferentiering proces opstår, når en celle bliver mere specialiseret til at udvikle sig til en flercellet organisme. Ændringer i størrelse, form, metaboliske aktiviteter og lydhørhed kan forekomme som følge af justeringer i genekspression. Celle mangfoldighed og differentiering, i denne art, involverer beslutninger truffet fra celle-celle interaktioner i veje til enten stilkceller eller sporeceller. Disse celleskæbner afhænger af deres miljø og mønsterdannelse. Derfor er organismen en glimrende model til at studere celledifferentiering.

Chemotaxis er defineret som en passage af en organisme mod eller væk fra en kemisk stimulus langs en kemisk koncentrationsgradient. Visse organismer demonstrerer kemotaksi, når de bevæger sig mod tilførsel af næringsstoffer. I D. discoideum udskiller amøben signalet, cAMP, ud af cellen og tiltrækker andre amøber til at migrere mod kilden. Hver amøbe bevæger sig mod en central amøbe, den der uddeler den største mængde cAMP -sekret. Sekretion af cAMP udstilles derefter af alle amøber og er en opfordring til dem om at begynde aggregering. Disse kemiske emissioner og amøbe -bevægelse forekommer hvert sjette minut. Amøberne bevæger sig mod koncentrationsgradienten i 60 sekunder og stopper, indtil den næste sekretion sendes ud. Denne adfærd hos individuelle celler har en tendens til at forårsage svingninger i en gruppe af celler, og kemiske bølger med varierende cAMP -koncentration formerer sig gennem gruppen i spiraler.
Et elegant sæt matematiske ligninger, der gengiver spiraler og streamingmønstre af D. discoideum, blev opdaget af matematiske biologer Thomas Höfer og Martin Boerlijst. Matematisk biolog Cornelis J. Weijer har bevist, at lignende ligninger kan modellere dens bevægelse. Ligningerne for disse mønstre påvirkes hovedsageligt af amoebebefolkningens tæthed, produktionen af cyklisk AMP og individuelle amøberes følsomhed over for cyklisk AMP. Det spiralformede mønster dannes af amøber i midten af en koloni, der roterer, når de sender bølger af cyklisk AMP ud.
Anvendelsen af cAMP som kemotaktisk middel er ikke etableret i nogen anden organisme. I udviklingsbiologi er dette et af de forståelige eksempler på kemotaksi, hvilket er vigtigt for en forståelse af menneskelig betændelse, gigt, astma, lymfocythandel og axonvejledning. Fagocytose bruges til immunovervågning og antigenpræsentation, mens celletypebestemmelse, cellesortering og mønsterdannelse er grundlæggende træk ved embryogenese, der kan studeres med disse organismer.
Bemærk dog, at CAMP -svingninger muligvis ikke er nødvendige for den kollektive cellemigration på flercellede stadier. En undersøgelse har fundet ud af, at cAMP-medieret signalering ændrer sig fra formerende bølger til en stabil tilstand på et multicellulært stadie af D. discoideum.
Thermotaxis er bevægelse langs en gradient af temperatur. Sneglene har vist sig at vandre langs ekstremt lavvandede gradienter på kun 0,05 ° C/cm, men den valgte retning er kompliceret; det ser ud til at være væk fra en temperatur omkring 2 ° C under den temperatur, de var blevet akklimatiseret til. Denne komplicerede adfærd er blevet analyseret ved computermodellering af adfærden og det periodiske mønster af temperaturændringer i jord forårsaget af daglige ændringer i lufttemperatur. Konklusionen er, at adfærden flytter snegle et par centimeter under jordoverfladen op til overfladen. Dette er en forbløffende sofistikeret adfærd fra en primitiv organisme uden tilsyneladende tyngdekraft.
Apoptose (programmeret celledød) er en normal del af artsudviklingen. Apoptose er nødvendig for korrekt afstand og udformning af komplekse organer. Omkring 20% af cellerne i D. discoideum ofrer sig altruistisk i dannelsen af den modne frugtlegeme. Under pseudoplasmodium (slug eller grex) fase af sin livscyklus har organismen dannet tre hovedtyper af celler: prestalk, prespore og anteriorlignende celler. Under kulminationen udskiller prestalk -cellerne et celluloselag og strækker sig som et rør gennem grexen. Efterhånden som de adskiller sig, danner de vakuoler og forstørrer og løfter de forspore celler. Stængelcellerne undergår apoptose og dør, da presporecellerne løftes højt over substratet. Presporecellerne bliver derefter til sporeceller, der hver bliver en ny myxamoeba ved spredning. Dette er et eksempel på, hvordan apoptose bruges til dannelse af et reproduktivt organ, den modne frugtkrop.
Et nylig stort bidrag fra Dictyostelium -forskning er kommet fra nye teknikker, der gør det muligt at visualisere aktiviteten af individuelle gener i levende celler. Dette har vist, at transkription forekommer i "burst" eller "pulser" ( transkriptionel bursting ) frem for at følge simpel probabilistisk eller kontinuerlig adfærd. Sprængende transkription ser nu ud til at være bevaret mellem bakterier og mennesker. Et andet bemærkelsesværdigt træk ved organismen er, at den har sæt af DNA -reparationsenzymer, der findes i humane celler, som mangler fra mange andre populære metazoanske modelsystemer. Defekter i DNA -reparation fører til ødelæggende menneskelige kræftformer, så evnen til at studere humane reparationsproteiner i en simpel transporterbar model vil vise sig uvurderlig.
Lab dyrkning
Denne organismes evne til let at blive dyrket i laboratoriet øger sin appel som modelorganisme. Mens D. discoideum kan dyrkes i flydende kultur, er det normalt dyrkes i petriskåle indeholdende næringsstof agar og overfladerne holdes fugtig. Kulturer vokser bedst ved 22–24 ° C (stuetemperatur). D. discoideum lever primært af E. coli , hvilket er tilstrækkeligt til alle faser af livscyklussen. Når fødevareforsyningen reduceres, aggregerer myxamoebæerne for at danne pseudoplasmodia. Snart er retten dækket med forskellige stadier af livscyklussen. Kontrol af fadet giver ofte mulighed for detaljerede observationer af udviklingen. Cellerne kan høstes på ethvert udviklingsstadium og dyrkes hurtigt.
Mens man dyrker D. discoidium i et laboratorium, er det vigtigt at tage hensyn til dets adfærdsmæssige reaktioner. For eksempel har den en affinitet over for lys, højere temperaturer, høj luftfugtighed, lave ionkoncentrationer og den sure side af pH -gradienten. Der udføres ofte eksperimenter for at se, hvordan manipulationer af disse parametre forhindrer, stopper eller fremskynder udviklingen. Variationer af disse parametre kan ændre hastigheden og levedygtigheden af kulturvækst. Frugtlegemerne, da det er det højeste udviklingsstadium, reagerer også meget på luftstrømme og fysiske stimuli. Det vides ikke, om der er en stimulus forbundet med frigivelse af sporer.
Undersøgelser af proteinekspression
Detaljeret analyse af proteinekspression i Dictyostelium er blevet hæmmet af store skift i proteinekspressionsprofilen mellem forskellige udviklingstrin og en generel mangel på kommercielt tilgængelige antistoffer for Dictyostelium -antigener. I 2013 rapporterede en gruppe ved Beatson West of Scotland Cancer Center om en antistoffri proteinvisualiseringsstandard for immunoblotting baseret på påvisning af MCCC1 ved hjælp af streptavidinkonjugater .
Legionærsygdom
Bakterieslægten Legionella omfatter den art, der forårsager legionærsygdom hos mennesker. D. discoideum er også vært for Legionella og er en egnet model til undersøgelse af infektionsprocessen. Specifikt deler D. discoideum med pattedyrsværtsceller et lignende cytoskelet og cellulære processer, der er relevante for Legionella -infektion, herunder fagocytose , membranhandel, endocytose , vesikelsortering og kemotaksi.
"Landbrug"
En rapport fra Nature i 2011 offentliggjorde fund, der demonstrerede en "primitiv landbrugsadfærd" i D. discoideum -kolonier. Beskrevet som en " symbiose " mellem D. discoideum og bakterielt bytte, var omkring en tredjedel af vildtindsamlede D. discoideum- kolonier engageret i bakteriens " husdyrbrug ", da bakterierne blev inkluderet i slimformens frugtlegemer. Indarbejdelsen af bakterierne i frugtlegemerne tillader "såning" af fødekilden ved sporespredningen, hvilket er særligt værdifuldt, hvis det nye sted er lavt i fødevareressourcer. Kolonier produceret fra "landbrugs" sporer viser typisk også den samme adfærd ved sporulering. Denne inkorporering har en omkostning forbundet med sig: De kolonier, der ikke spiser alle byttedyrbakterierne, producerer mindre sporer, der ikke kan sprede sig så bredt. Derudover findes der langt mindre fordel for bakterier, der indeholder sporer, der lander i en fødevarerig region. Denne balance mellem omkostninger og fordele ved adfærden kan bidrage til, at et mindretal af D. discoideum -kolonier deltager i denne praksis.
D. discoideum er kendt for at spise gram-positive såvel som gram-negative bakterier , men nogle af de fagocytiserede bakterier, herunder nogle menneskelige patogener, er i stand til at leve i amøberne og forlade uden at dræbe cellen. Hvornår de kommer ind i cellen, hvor de bor, og når de forlader cellen, kendes ikke. Forskningen er endnu ikke afgørende, men det er muligt at tegne en generel livscyklus for D. discoideum tilpasset landmandskloner for bedre at forstå denne symbiotiske proces.

På billedet kan man se de forskellige stadier. For det første lukkes bakterier i sultestadiet inden for D. discoideum , efter indtræden i amøber, i et fagosom blokeres fusionen med lysosomer, og disse umodnede fagosomer er omgivet af værtscelleorganeller såsom mitokondrier, vesikler og en flerlags membran afledt fra den ru endoplasmatiske retikulum (RER) af amøber. RER's rolle i den intracellulære infektion kendes ikke, men RER er ikke påkrævet som proteinkilde for bakterierne. Bakterierne opholder sig inden for disse fagosomer under aggregeringen og de multicellulære udviklingstrin. Amøberne bevarer deres individualitet, og hver amøbe har sin egen bakterie. Under kulminationstrinnet, når sporer produceres, passerer bakterierne fra cellen til sorus ved hjælp af en cytoskeletal struktur, der forhindrer ødelæggelse af værtsceller. Nogle resultater tyder på, at bakterierne udnytter eksocytosen uden at dræbe cellen. Fritlevende amøber synes at spille en afgørende rolle for vedholdenhed og spredning af nogle patogener i miljøet. Forbigående forbindelse med amøber er blevet rapporteret for en række forskellige bakterier, herunder Legionella pneumophila , mange Mycobacterium -arter, Francisella tularensis og Escherichia coli , blandt andre. Landbrug ser ud til at spille en afgørende rolle for patogeners overlevelse, da de kan leve og replikere inde i D. discoideum , hvilket gør husdyrbrug. Naturens rapport har gjort et vigtigt fremskridt i kendskabet til amøbisk adfærd, og den berømte spanske sætning oversat med “du er mere dum end en amøbe” mister fornuften, fordi amøber er et glimrende eksempel på social adfærd med en fantastisk koordination og fornemmelse af ofre til gavn for arten.
Sentinel celler
Sentinelceller i Dictyostelium discoideum er fagocytiske celler, der er ansvarlige for at fjerne giftigt materiale fra sneglefasen i den sociale cyklus. Generelt runde i form, er disse celler til stede i sneglekappen, hvor det viser sig, at de cirkulerer frit. Afgiftningsprocessen opstår, når disse celler opsluger toksiner og patogener i sneglen gennem fagocytose . Derefter klumper cellerne sig i grupper på fem til ti celler, som derefter fastgøres til sneglens indre kappe. Skeden slæbes af, når sneglen migrerer til et nyt sted på jagt efter madbakterier.
Sentinelceller udgør cirka 1% af det samlede antal slugceller, og antallet af sentinelceller forbliver konstant, selvom de frigives. Dette indikerer en konstant regenerering af sentinelceller i sneglene, når de fjernes sammen med toksiner og patogener. Sentinelceller er til stede i sneglen, selv når der ikke er toksiner eller patogener, der skal fjernes. Sentinelceller har været lokaliseret i fem andre arter af Dictyostelia , hvilket tyder på, at sentinelceller kan beskrives som en generel egenskab ved det medfødte immunsystem i sociale amøber.
Virkninger af landbrugsstatus på sentinelceller
Antallet af vagtpostceller varierer afhængigt af landbrugsstatus for vild D. discoideum . Når de blev udsat for et giftigt miljø skabt ved brug af ethidiumbromid, blev det vist, at antallet af sentinelceller pr. Millimeter var lavere for landmænd end ikke-landmænd. Dette blev afsluttet ved at observere de stier, der blev efterladt, da sneglene migrerede og tælle antallet af sentinelceller til stede i en millimeter. Antallet af vagtpostceller påvirker imidlertid ikke sporeproduktionen og levedygtigheden hos landmænd. Landmænd udsat for et giftigt miljø producerer det samme antal sporer som landmænd i et giftfrit miljø, og sporbarheden var den samme mellem landmændene og ikke-landmændene. Når Clade 2 Burkholderia eller landmandsassocierede bakterier fjernes fra landmændene, lignede sporproduktion og levedygtighed ikke-landmændenes. Det foreslås således, at bakterier båret af landmændene yder en ekstra rolle for landmændene mod potentiel skade på grund af toksiner eller patogener.
Klassificering og fylogeni
I ældre klassifikationer blev Dictyostelium placeret i den nedlagte polyfyletiske klasse Acrasiomycetes . Dette var en klasse af cellulære slimforme, som var præget af sammenlægning af individuelle amøber til en flercellet frugtlegeme, hvilket gjorde det til en vigtig faktor, der relaterede acrasiderne til dictyosteliderne.
Nyere genomiske undersøgelser har vist, at Dictyostelium har opretholdt mere af sin forfædres genomdiversitet end planter og dyr, selvom proteombaseret fylogeni bekræfter, at amoebozoer afviger fra slægten mellem dyr og svampe efter splittelsen mellem planter og dyr. Underklasse Dictyosteliidae, ordre Dictyosteliales er en monofyletisk samling inden for Mycetozoa, en gruppe, der omfatter protostelid-, dictyostelid- og myxogastrid -slimformene. Forlængelsesfaktor-1α (EF-1α) dataanalyser understøtter Mycetozoa som en monofyletisk gruppe, selvom rRNA-træer placerer det som en polyfyletisk gruppe. Disse data understøtter endvidere tanken om, at dictyostelid og myxogastrid er tættere forbundet med hinanden, end de er protosteliderne. EF-1α-analyse placerede også Mycetozoa som den umiddelbare udgruppe for dyresvampekladen. Seneste fylogenetiske data placerer dictyostelider fast i supergruppen Amoebozoa sammen med myxomycetes . I mellemtiden har protostelider vist sig at være polyfyletiske, deres stilkede frugtlegemer er et konvergent træk ved flere ikke -relaterede slægter.
Genom
Den D. discoideum genom sekventering Projektet blev afsluttet og offentliggjort i 2005 af et internationalt samarbejde med institutter. Dette var det første fritlevende protozo-genom, der blev fuldstændigt sekvenseret. D. discoideum består af et 34-Mb haploid genom med en basesammensætning på 77% [A+T] og indeholder seks kromosomer, der koder for omkring 12.500 proteiner. Sekventering af D. discoideum genomet giver en mere indviklet undersøgelse af dens cellulære og udviklingsmæssige biologi.
Tandem -gentagelser af trinucleotider er meget rigelige i dette genom; en klasse af genomet er samlet, hvilket får forskere til at tro, at det fungerer som centromerer. Gentagelserne svarer til gentagne sekvenser af aminosyrer og menes at blive ekspanderet ved nukleotidudvidelse. Udvidelse af trinucleotid -gentagelser forekommer også hos mennesker, hvilket generelt fører til mange sygdomme. At lære, hvordan D. discoideum -celler udholder disse aminosyre -gentagelser, kan give indsigt, så mennesker kan tolerere dem.
Hvert genom sekvenseret genom spiller en vigtig rolle i identifikationen af gener, der er blevet vundet eller tabt over tid. Sammenlignende genomiske undersøgelser tillader sammenligning af eukaryote genomer. En fylogeni baseret på proteomet viste, at amoebozoa afvigede fra dyr-svampeafstamning efter plant-dyr-splittelsen. The D. discoideum -genomet er bemærkelsesværdigt, fordi dens mange kodede proteiner er almindeligt forekommende i svampe, planter og dyr.
Databaser
- DictyBase - generel genomisk database om Dictyostelium discoideum
- Membranome-database giver information om single-pass transmembranproteiner fra Dictyostelium og flere andre organismer
Referencer
Yderligere læsning
- Mary S. Tyler (2000). Developmental Biology: A Guide for Experimental Study . 2. udg . Sinauer Associates. s. 31–34. ISBN 978-0-87893-843-8.
- Scott F. Gilbert (2006). Udviklingsbiologi. 8. udg . Sinauer. s. 36–39. ISBN 978-0-87893-250-4.