
Opabinia -Opabinia
|
Opabinia |
|
|---|---|

|
|
| Fossil prøve ( lektotype ) udstillet på Smithsonian i Washington, DC | |
|
Videnskabelig klassificering |
|
| Kongerige: | Animalia |
| Phylum: | Arthropoda |
| Klasse: | † Dinocaridida |
| Familie: | † Opabiniidae |
| Slægt: |
† Opabinia Walcott , 1912 |
| Arter: |
† O. regalis
|
| Binomisk navn | |
|
† Opabinia regalis
Walcott , 1912
|
|
Opabinia regalis er en uddød , stamceller gruppe leddyr fundet i Mellemøsten Kambrium Burgess Shale Lagerstätte (505 millioner år siden) af British Columbia . Opabinia var et blødt dyr med en længde på op til 7 cm, og dets segmenterede bagagerum havde klapper langs siderne og en ventilatorformet hale. Hovedet viser usædvanlige træk: fem øjne , en mund under hovedet og bagudvendt og en kløet snabel, der sandsynligvis førte mad til munden. Opabinia levede sandsynligvis på havbunden og brugte snabel til at opsøge små, bløde fødevarer. Færre end tyve gode eksemplarer er blevet beskrevet; 3 eksemplarer af Opabinia kendes fra Greater Phyllopod -sengen , hvor de udgør mindre end 0,1% af samfundet.
Da den første grundige undersøgelse af Opabinia i 1975 afslørede dens usædvanlige træk, blev det antaget at være uden relation til nogen kendt filum eller måske en slægtning til leddyr og annelid -forfædre . Senere undersøgelser siden slutningen af 1990'erne understøtter imidlertid konsekvent dets affinitet som medlem af basale leddyr sammen med de nært beslægtede radiodonts ( Anomalocaris og slægtninge) og forgyldte lobopodianer ( Kerygmachela og Pambdelurion ).
I 1970'erne var der en løbende debat om, hvorvidt flercellede dyr pludselig dukkede op under den tidlige kambriske, i en begivenhed kaldet den kambriske eksplosion , eller var opstået tidligere, men uden at efterlade fossiler. I første omgang blev Opabinia betragtet som et stærkt bevis for den "eksplosive" hypotese. Senere opdagelsen af en hel række lignende lobopodian dyr, nogle med tættere ligheder med leddyr, og udvikling af ideen om stamceller grupper foreslået, at den tidlige Kambrium var en tid med relativt hurtigt evolution , men en, der kunne forstås uden at antage nogen entydig evolutionære processer.
Opdagelseshistorie
Charles Doolittle Walcott fandt i Burgess -skiferen ni næsten fuldstændige fossiler af Opabinia regalis og et par af det, han klassificerede som Opabinia? medier og offentliggjorde en beskrivelse af alle disse i 1912. Det generiske navn stammer fra Opabin -passet mellem Mount Hungabee og Mount Biddle , sydøst for Lake O'Hara , British Columbia, Canada . I 1966–1967 fandt Harry B. Whittington endnu et godt eksemplar, og i 1975 offentliggjorde han en detaljeret beskrivelse baseret på meget grundig dissektion af nogle eksemplarer og fotografier af disse eksemplarer tændt fra forskellige vinkler. Whittingtons analyse dækkede ikke Opabinia? medier : Walcotts eksemplarer af denne art kunne ikke identificeres i hans samling. I 1960 beskrev russiske paleontologer eksemplarer, de fandt i Norilsky -regionen i Sibirien og betegnede Opabinia norilica , men disse fossiler var dårligt bevarede, og Whittington følte ikke, at de gav nok information til at blive klassificeret som medlemmer af slægten Opabinia .
Hændelse
Alle de anerkendte Opabinia -eksemplarer, der hidtil er fundet, stammer fra " Phyllopod -sengen " i Burgess -skiferen i Canadian Rockies i British Columbia.
I 1997 rapporterede Briggs og Nedin fra South Australia Emu Bay Shale om et nyt eksemplar af Myoscolex, der var meget bedre bevaret end tidligere prøver, hvilket fik dem til at konkludere, at det var en nær slægtning til Opabinia - selvom denne fortolkning senere blev stillet spørgsmålstegn ved Dzik, som konkluderede i stedet, at Myoscolex var en annelid orm.
Morfologi
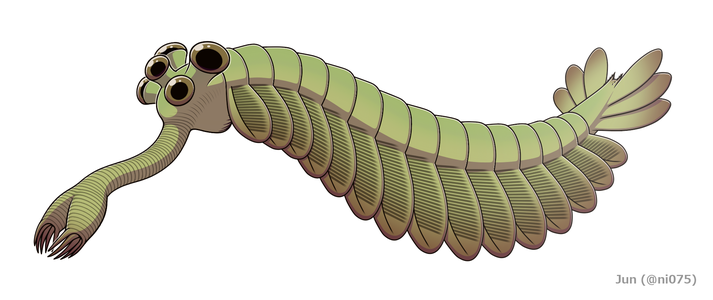
Restaurering

Størrelsesvurdering



Opabinia så så mærkelig ud, at publikum ved den første præsentation af Whittingtons analyse lo. Længden af Opabinia regalis fra hoved (undtagen snabel) til haleenden varierede mellem 4 centimeter (1,6 tommer) og 7 centimeter (2,8 tommer). En af de mest karakteristiske karakterer ved Opabinia er den hule snabel , hvis samlede længde var omkring en tredjedel af kroppens og projiceret ned fra under hovedet. Snablen blev stribet som en støvsugerslange og fleksibel, og den endte med en klo-lignende struktur, hvis terminal kanter bar 5 rygsøjler, der ragede indad og fremad. Den bilaterale symmetri og laterale (i stedet for lodrette som rekonstrueret af Whittington 1975) anordning af kloen tyder på, at den repræsenterer et par sammensmeltede frontale vedhæng, der kan sammenlignes med radiodonts og forgyldte lobopodians . Hovedet bar fem stilkede øjne: to nær fronten og temmelig tæt på midten af hovedet, pegende opad og fremad; to større øjne med længere stilke nær hovedets bageste og ydre kanter, pegende opad og sidelæns; og et enkelt øje mellem det større par stilkede øjne, der peger opad. Det er blevet antaget, at øjnene alle var sammensatte , ligesom andre leddyrs laterale øjne, men denne rekonstruktion, som ikke understøttes af noget bevis, er "noget fantasifuld". Munden var under hovedet, bag sneglen og pegede baglæns , så fordøjelseskanalen dannede en U-bøjning på vej mod dyrets bagside. Snablen syntes tilstrækkelig lang og fleksibel til at nå munden.
Hoveddelen af kroppen var typisk omkring 5 millimeter (0,20 tommer) bred og havde 15 segmenter, på hvilke der hver var par flapper (lober), der pegede nedad og udad. Klapperne overlappede, så forsiden af hver var dækket af bagkanten af den foran den. Kroppen sluttede med, hvad der lignede et enkelt konisk segment med tre par overlappende haleblade, der pegede op og ud og dannede en hale som en V-formet dobbelt ventilator.


Forskellige fortolkninger af klapper og gillestrukturer i Opabinia regalis
A: Whittington (1975), B: Bergström (1986), C: Budd (1996), D: Zhang & Briggs (2007), E: Budd & Daley (2011)
Opabinia -tværsnit baseret på Budd og Daley (2011)


Fortolkninger af andre træk ved Opabinia -fossiler er forskellige. Da dyrene ikke havde mineraliseret rustning eller endda hårde organiske eksoskeletoner som andre leddyr, blev deres kroppe fladtrykt, da de blev begravet og fossiliseret, og mindre eller indre træk fremstår som markeringer inden for fossilernes konturer.
Whittington (1975) fortolkede gællerne som parrede forlængelser fastgjort dorsalt til baserne på alle undtagen de første klapper på hver side og troede, at disse gæller var flade nedenunder, havde overlappende lag ovenpå. Bergström (1986) afslørede, at "overlappende læg" var rækker af individuelle klinger, tolket flapperne som en del af dorsale belægninger ( tergit ) over kroppens øvre overflade, med knive fastgjort under hver af dem. Budd (1996) troede, at gællebladene var fastgjort langs forkanterne på dorsalsiden af alle undtagen de første klapper. Han fandt også mærker inde i flappernes forreste kanter, som han fortolkede som interne kanaler, der forbinder gællerne med det indre af kroppen, ligesom Whittington fortolkede mærket langs snablen som en intern kanal. Zhang og Briggs (2007) fortolkede imidlertid, at alle klapper har posterior afstand, hvor gyllebladene er fastgjort. Budd og Daley (2011) afviser rekonstruktionen af Zhang & Briggs, idet klapperne har komplette posteroir kanter som tidligere rekonstruktioner. De følger for det meste rekonstruktionen af Budd (1996) med ændringer på nogle detaljer (f.eks. Har det første klappepar også gæller; fastgørelsespunktet for gælleblade placeret mere bagud end tidligere).


Whittington (1975) fandt tegn på næsten trekantede træk langs kroppen og konkluderede, at de var interne strukturer, sandsynligvis sidelæns forlængelser af tarmen ( diverticula ). Chen et al. (1994) fortolkede dem som indeholdt i lober langs siderne. Budd (1996) troede "trekanter" var for bred til at passe inden Opabinia ' s slank krop, og at tværsnit synspunkter viste de blev fæstnet separat fra og sænk end lapperne, og udvides under kroppen. Han fandt senere eksemplarer, der syntes at bevare benenes ydre neglebånd. Han fortolkede derfor "trekanterne" som korte, kødfulde, koniske ben (lobopoder). Han fandt også små mineraliserede pletter på spidserne af nogle og fortolkede disse som kløer. Under denne rekonstruktion blev gællebærende klap og lobopod homologiseret til den ydre gillegren og indre bengren af leddyr biramøse lemmer set i Marrella , trilobitter og krebsdyr . Zhang og Briggs (2007) analyserede den "trekantes" kemiske sammensætning og konkluderede, at de havde samme sammensætning som tarmen, og var derfor enige med Whittington om, at de var en del af fordøjelsessystemet. I stedet betragtes Opabinia ' s lap + gill arrangement som en tidlig form af leddyr lemmer før det opdeles i en biramous struktur. Denne lignende kemiske sammensætning er imidlertid ikke kun forbundet med fordøjelseskanalen; Budd og Daley (2011) antyder, at det repræsenterer mineralisering, der dannes inden for væskefyldte hulrum i kroppen, hvilket er i overensstemmelse med hule lobopoder set i utvetydige lobopodiske fossiler. De præciserer også, at tarmdivertikulaen i Opabinia er en række cirkulære tarmkirtler individualiseret fra "trekanterne". Mens de blev enige om fraværet af terminalkløer, er tilstedeværelsen af lobopoder i Opabinia stadig en sandsynlig fortolkning.
Livsstil
Den måde, hvorpå Burgess-skiferdyrene blev begravet, ved et mudderskred eller en sedimentbelastet strøm, der fungerede som sandstorm, tyder på, at de levede på havbundens overflade. Opabinia brugte sandsynligvis sin snabel til at søge sedimentet efter madpartikler og sende dem til munden. Da der ikke er tegn på noget, der kan fungere som kæber, var dets mad formodentlig lille og blød. Den parrede tarm diverticula kan øge effektiviteten af madfordøjelse og indtagelse af ernæring. Whittington (1975) troede på, at Opabinia ikke havde nogen ben, troede, at den kravlede på dens lapper, og at den også kunne have svømmet langsomt ved at klappe lapperne, især hvis den timede bevægelserne til at skabe en bølge med metakoral bevægelse af dens lober. På den anden side syntes han, at kroppen ikke var fleksibel nok til at tillade fisklignende bølger af hele kroppen.
Klassifikation
|
|||||||||||||||||||||||||||||||||||||||
| Opsummeret fylogeni mellem Opabinia og andre Ecdysozoan taxa. |





I betragtning af hvordan paleontologers rekonstruktioner af Opabinia adskiller sig, er det ikke overraskende, at dyrets klassifikation er stærkt debatteret i løbet af 20 århundrede. Charles Doolittle Walcott , den originale beskrivelse , betragtede det som et anostrakansk krebsdyr i 1912. Idéen blev fulgt af G. Evelyn Hutchinson i 1930, der gav den første rekonstruktion af Opabinia som en anostracan, der svømmede på hovedet. Alberto Simonetta leverede en ny rekonstruktion af Opabinia i 1970 meget forskellig fra Hutchinson's, med masser af leddyrsfunktioner (f.eks. Dorsal eksoskelet og leddede lemmer), der minder om Yohoia og Leanchoilia . Leif Størmer , efter tidligere arbejde af Percy Raymond , mente, at Opabinia tilhørte de såkaldte "trilobitoider" ( trilobitter og lignende taxa). Efter hans grundige analyse konkluderede Harry B. Whittington , at Opabinia ikke var leddyr i 1975, da han ikke fandt beviser for leddyr leddede led, og intet lignede den fleksible, sandsynligvis væskefyldte snabel ved leddyr. Selv om han forlod Opabinia ' s klassifikation over familiens niveau åben, den annuleret, men ikke leddelte krop og de usædvanlige laterale flapper med gæller overtalte ham, at det kan have været en repræsentant for det fædrene bestand fra oprindelsen af ledorme og leddyr, to forskellige dyr phyla (henholdsvis Lophotrochozoan og Ecdysozoan ), som på det tidspunkt stadig blev anset for at være nære slægtninge (forenet under Articulata ).
I 1985 udgav Derek Briggs og Whittington en større omskrift af Anomalocaris , også fra Burgess Shale. Kort tid efter bemærkede den svenske paleontolog Jan Bergström i 1986 om ligheden mellem Anomalocaris og Opabinia , foreslog, at de to dyr var beslægtede, da de delte adskillige træk (f.eks. Sideklapper , gylleblade , stilkede øjne og specialiserede frontal vedhæftede filer). Han klassificerede dem som primitive leddyr, selvom han mente, at leddyr ikke er et enkelt fylum .
I 1996 fandt Graham Budd , hvad han betragtede som tegn på korte ben uden led i Opabinia . Hans undersøgelse af den forgyldte lobopodian Kerygmachela fra Sirius Passet lagerstätte , for cirka 518 millioner år siden og over 10 millioner år ældre end Burgess -skiferen, overbeviste ham om, at denne prøve havde lignende ben. Han betragtede benene på disse to slægter meget lig dem hos Burgess Shale lobopodian Aysheaia og de moderne onychophorans ( fløjlsorm ), der betragtes som bærere af talrige ansektrale træk, der deles af forfædrene med leddyr. Efter at have undersøgt flere sæt af funktioner, der deles af disse og lignende lobopodianm trak han op en "bred skala rekonstruktion af leddyr stilk-gruppe ", med andre ord leddyr og hvad han anså for at være deres evolutionære basale medlemmer. Et slående træk ved dette familietræ er, at moderne tardigrades (vand bjørne) kan være Opabinia ' s nærmeste levende evolutionære slægtninge. På den anden side har Hou et al. (1995, 2006) snarere foreslået, at Opabinia er medlem af usædvanlige cykloneuraliske orme med konvergerende leddyrsfunktioner .
Selvom Zhang og Briggs (2007) var uenige med Budd diagnose at Opabinia ' s 'trekanter' var ben, ligheden de så mellem Opabinia ' s lap + gælle arrangement og leddyr biramous lemmer førte dem til at konkludere, at Opabinia var meget tæt knyttet til leddyr . Faktisk præsenterede de et slægtstræ, der meget lignede Budds, bortset fra at deres ikke nævnte tardigrader. Uanset de forskellige morfologiske fortolkninger konkluderede alle større undersøgelser siden 1980'erne på samme måde, at ligheden mellem Opabinia og leddyr (f.eks. Stalkede øjne; dorsal segmentering; posterior mouth; fused appendages; gill-like lem grenene) er taksonomisk signifikante.
Siden 2010'erne understøttes det foreslåede tætte forhold mellem Opabinia og tardigrader/cycloneuralians ikke længere, mens Opabinias affinitet som stammegruppe leddyr sammen med Radiodonta (en klade, der omfatter Anomalocaris og dets slægtninge) og forgyldte lobopodianer er bredt accepteret, blev konsekvent genoprettet ved multipel fylogenetisk analyse, samt nye opdagelser såsom tilstedeværelsen af leddyrlignende tarmkirtler og mellemliggende taxon Kylinxia .
Teoretisk betydning

Opabinia gjorde det klart, hvor lidt man vidste om bløde dyr, som normalt ikke efterlader fossiler. Da Whittington beskrev det i midten af 1970'erne, var der allerede en kraftig debat om den tidlige udvikling af dyr . Preston Cloud argumenterede i 1948 og 1968 for, at processen var "eksplosiv", og i begyndelsen af 1970'erne udviklede Niles Eldredge og Stephen Jay Gould deres teori om punkteret ligevægt , der betragter evolution som lange intervaller af nær-stasis "punkteret" af korte perioder med hurtig forandring. På den anden side argumenterede Wyatt Durham og Martin Glaessner på samme tid begge for, at dyreriget havde en lang proterozoisk historie, der var skjult af manglen på fossiler. Whittington (1975) konkluderede, at Opabinia og andre taxaer såsom Marrella og Yohoia ikke kan rummes i moderne grupper. Dette var en af de primære årsager til, at Gould i sin bog om Burgess -skiferen , Wonderful Life , mente, at det tidlige kambriumske liv var meget mere mangfoldigt og "eksperimentelt" end noget senere sæt dyr, og at den kambriske eksplosion var en virkelig dramatisk begivenhed, muligvis drevet af usædvanlige evolutionære mekanismer. Han betragtede Opabinia som så vigtig for at forstå dette fænomen, at han ønskede at kalde sin bog Hyldest til Opabinia .
Imidlertid fulgte snart andre opdagelser og analyser, der afslørede lignende dyr som Anomalocaris fra Burgess- skiferen og Kerygmachela fra Sirius Passet . Et andet Burgess Shale -dyr , Aysheaia , blev betragtet som meget ligner moderne Onychophora , der betragtes som nære slægtninge til leddyr. Paleontologer definerede en gruppe kaldet lobopodianer til at omfatte fossile panarthropoder, der menes at være nære slægtninge til onychophoraner, tardigrader og leddyr, men mangler leddede lemmer. Denne gruppe blev senere bredt accepteret som en parafyletisk kvalitet, der førte til oprindelsen af eksisterende panarthropod phyla.

Mens denne diskussion om specifikke fossiler såsom Opabinia og Anomalocaris foregik i slutningen af 20-tallet, begrebet stamceller grupper blev indført for at dække evolutionære "tanter" og "fætre". En krongruppe er en gruppe af nært beslægtede levende dyr plus deres sidste fælles forfader plus alle dens efterkommere. En stamgruppe indeholder udløbere fra medlemmer af slægten tidligere end krongruppens sidste fælles forfader; det er et relativt begreb, for eksempel er tardigrader levende dyr, der udgør en krongruppe i sig selv, men Budd (1996) betragtede dem også som værende en stamgruppe i forhold til leddyrene. At se på mærkelige organismer som Opabinia på denne måde gør det muligt at se, at mens den kambriske eksplosion var usædvanlig, kan den forstås ud fra normale evolutionære processer.
Se også
- Kropsplan - Sæt af morfologiske træk, der er fælles for medlemmer af et dyrefyl
- Radiodonta - Uddød rækkefølge af kambriske leddyr
Referencer
Yderligere læsning
- Bergström, J. (1987). "The Cambrian Opabinia and Anomalocaris ". Lethaia . 20 (2): 187–188. doi : 10.1111/j.1502-3931.1987.tb02037.x .
- Briggs, DEG; Whittington, HB (1987). "Affiniteterne for de kambriske dyr Anomalocaris og Opabinia ". Lethaia . 20 (2): 185–186. doi : 10.1111/j.1502-3931.1987.tb02036.x .
eksterne links
- " Opabinia regalis " . Burgess Shale Fossil Gallery . Virtuelt museum i Canada. 2011.
- Smithsonian -side om Opabinia , med foto af Burgess Shale -fossil