
Radiodonta -Radiodonta
| Radiodonta |
|
|---|---|

|
|
| Venstre mod højre, top til bund: Amplectobelua symbrachiata , Anomalocaris canadensis , Aegirocassis benmoulai , Peytoia nathorsti , Lyrarapax unguispinus , Cambroraster falcatus og Hurdia victoria | |
| Videnskabelig klassifikation | |
| Kongerige: | |
| Fylde: | |
| Klasse: | |
| Bestille: |
† Radiodonta
Collins , 1996 |
| Familier | |
|
|
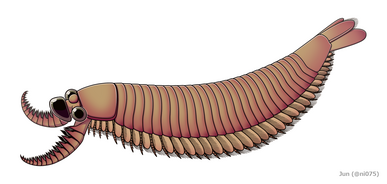
Radiodonta er en uddød orden af stammegruppe leddyr , der havde succes på verdensplan under den kambriske periode. De kan omtales som radiodonts , radiodontans , radiodontids , anomalocarids eller anomalocaridids , selvom de to sidste oprindeligt refererer til familien Anomalocarididae , som tidligere omfattede alle arter af denne orden, men nu er begrænset til kun nogle få arter. Radiodonts er kendetegnet ved deres karakteristiske frontale vedhæng, som er morfologisk forskelligartede og bruges til en række forskellige funktioner. Radiodonts omfattede de tidligste kendte store rovdyr, men de omfattede også sedimentsigter og filterfødere. Nogle af de mest kendte arter af radiodonter er den kambriske taxa Anomalocaris canadensis , Hurdia victoria , Peytoia nathorsti , Titanokorys gainessii , Cambroraster falcatus og Amplectobelua symbrachiata , den ordoviciske Aegirocassis benmoulai og den Devoniske Schinderhan .
Etymologi
Navnet Radiodonta (latin for radius "talte af et hjul" og græsk for odoús "tand") refererer til det radiale arrangement af tandplader (oral kegle), der omgiver munden, selvom disse træk foreslås at være fraværende hos nogle radiodont-arter.
Definition
Den oprindelige diagnose af orden Radiodonta i 1996 er som følger:
Radiodontids er bilateralt symmetriske, aflange leddyr med en ikke-mineraliseret kutikula, typisk mest robust i kæber og kløer. Kroppen er underopdelt i to tagmata , meget ligesom prosoma og opisthosoma hos chelicerede leddyr. Typisk viser den forreste del ingen ydre segmentering, bærer et par præorale kløer, et par fremtrædende øjne og ventrale kæber med udstrålende tænder. Nogle former har yderligere rækker af tænder og tre eller fire postorale gnathobasiske lemmerpar . Stammen er metamerisk , typisk med omkring 13 segmenter, der lateralt udvikler overlappende lapper til svømning og gæller til åndedræt, og kan ende i en fremtrædende tredelt hale. Nogle former har gnathobasiske stammelemmer.
I 2014 blev kladen Radiodonta defineret fylogenetisk som en klade inklusive enhver taxa tættere på Anomalocaris canadensis end Paralithodes camtschaticus . I 2019 blev det omdefineret morfologisk som dyrebærende hovedskjoldkompleks med centrale (H-) og laterale (P-) elementer; udvækster (enditter) fra frontale vedhæng, der bærer hjælperyg; og reducerede forreste flapper eller bånd af lameller (setalblade) og kraftig tilspidsning af kroppen fra anterior til posterior.
Beskrivelse

De fleste radiodonter var betydeligt større end andre kambriske faunaer , med typiske kropslængder varierende fra 30 til 50 centimeter. Den største beskrevne radiodont er Ordovician Aegirocassis benmoulai , som kan være blevet op til to meter lang. Et næsten komplet eksemplar af en juvenil Lyrarapax unguispinus målte kun 18 millimeter (0,71 tommer), hvilket gør den blandt de mindste kendte radiodontprøver, selvom voksne nåede en længde på 8 centimeter (3,1 tommer) Et isoleret frontalt vedhæng af en hurdiid med en længde mindre mere end halvdelen af den unge Lyrarapax er kendt, men det vides ikke, om dette eksemplar vedrører en voksen. Den største kendte kambriske radiodont var Laminacaris , selvom den kun er kendt fra frontale vedhæng, havde den en anslået kropslængde på op til 78,4 centimeter (30,9 in) baseret på Anomalocaris . Anomalocaris og Amplectobelua er også store, nåede 37,8 centimeter (14,9 tommer) og 48 centimeter (19 tommer) (der var et skøn, at Houcaris saron (tidligere Anomalocaris saron ) nåede 56 centimeter (22 tommer), men prøven blev brugt til at estimere kroppen længde hører ikke længere til den art); den kambriske hurdiid Titanokorys nærmede sig den i størrelse med en anslået kropslængde på cirka 50 centimeter (20 tommer).
Kroppen af en radiodont kan opdeles i to regioner: hoved og krop. Hovedet er sammensat af kun ét kropssegment kendt som den okulære somit, dækket af scleritter (hovedskjoldkompleks), med leddyrbehandlede frontale vedhæng, ventrale munddele (oral kegle) og stilkede sammensatte øjne . Den tilspidsede stamme er sammensat af flere kropssegmenter, der hver især er forbundet med par af klapper og gællelignende strukturer (setalblade).
Frontal vedhæng


De forreste strukturer på hovedet er et par frontale vedhæng, som er blevet omtalt som 'kløer', 'gribende vedhæng', 'fødevedhæng' eller 'store vedhæng' i tidligere undersøgelser (det sidste udtryk er frarådt, da homologien mellem frontale vedhæng og de originale, megacheirske store vedhæng er tvivlsomme.). De er sklerotiserede (hærdede) og arthropodiserede (segmenterede), og bærer ventrale enditter (rygge) på de fleste af deres podomerer (segmentale enheder), og enditterne kan bære yderligere rækker af hjælperygsøjler på deres for- og bagkant. Det frontale vedhæng består af to områder: skaftet ('peduncle', 'base' eller 'promixal region' i nogle undersøgelser) og den distale artikulerede region (også kaldet 'klo'). Et trekantet område dækket af bløde kutikula (arthrodial membran) kan forekomme på den ventrale side mellem podomerer og give fleksibilitet. Deres præ-okulære og protocerebrale oprindelse tyder på, at de er homologe med de primære antenner af Onychophora og labrum af Euarthropoda (alle opstod fra okulær somit ), og ikke homologe med chelicerae af Chelicerata eller antennerne eller ' store vedhæng ' af andre leddyr, som er deutocerebrale (opstået fra post-okulær somit 1). Da morfologien af frontalvedhængene, især rygsøjlens, altid er forskellig fra art til art, er det et af de vigtigste midler til artsidentifikation. Faktisk kendes mange radiodonter kun fra en håndfuld fossiliserede frontalvedhæng.

Frontale vedhæng af Anomalocarididae , Amplectobeluidae og muligvis beslægtede arter

Frontale vedhæng af Tamisiocarididae

Frontale vedhæng af Hurdiidae



Oral kegle

Munden er på den ventrale side af hovedet, bag fastgørelsespunktet for frontale vedhæng og er omgivet af en ring af tandplader, der danner munddelen kendt som oral kegle ('kæber' i tidligere undersøgelser). 3 eller 4 tandplader kan være forstørrede, hvilket giver den orale kegle et triradialt (f.eks . Anomalocaris ) eller tetraradialt (f.eks . Hurdiidae , Lyrarapax ) udseende. Den indvendige kant af tandpladerne har pigge, der vender mod mundåbningen. Yderligere rækker af indvendige tandplader kan forekomme i nogle hurdiid- slægter. Detaljerede rekonstruktioner af nogle amplektobeluiide orale kegler er spekulative, men de præsenterede muligvis ikke et typisk radialt arrangement.
Hovedskleritter, øjne og krop

Trehovedet sclerite ( carapace ) kompleks dannet af et centralt H-element (anterior sclerite eller hovedskjold) og et par P-elementer (laterale scleritter) dækker den dorsale og lateroventale overflade af dyrets hoved. P-elementerne kan forbindes med hinanden såvel som H-elementet med en smal anterior forlængelse (P-element-hals eller 'næb'). Hovedskleritterne er små og ægformede hos Anomalocarididae og Amplectobeluidae , men ofte forstørrede hos Hurdiidae , svarede til deres distinkte kropsformer (strømlinet hos Anomalocarididae/Amplectobeluidae, men ofte kompakte hos Hurdiidae). Hovedet bar to stilkede sammensatte øjne , som kan have haft mobilitet, og er placeret mellem hullerne dannet af de bageste områder af H-elementet og P-elementerne.


I modsætning til den oprindelige diagnose kan opdelingen af kropssegmenter (segmentale grænser) være synlige eksternt, og intet kendt medlem af Radiodonta (undtagen den formodede radiodont Cucumericrus ) er kendt for at have pediforme trunkvedhæng (ben). Trunken har adskillige kropssegmenter ( somitter ), som tilspidser fra anterior til posterior, med de forreste 3 eller 4 segmenter væsentligt indsnævret til en halsregion.

Variationer af radiodont kropsflapper
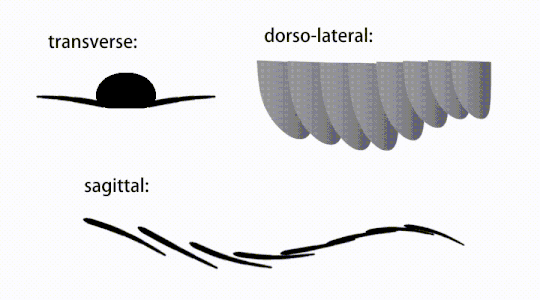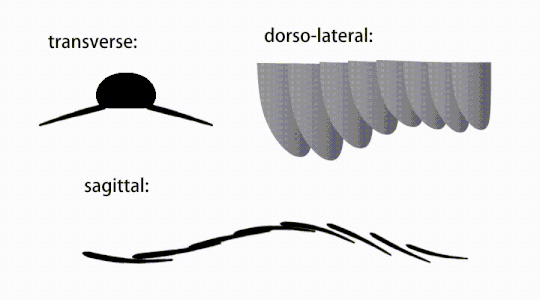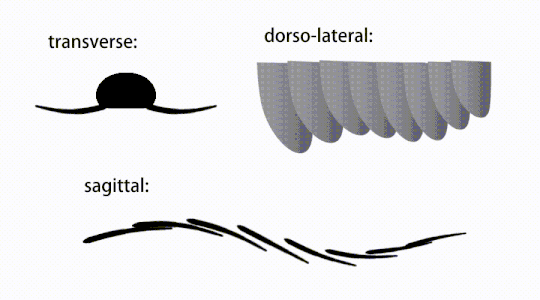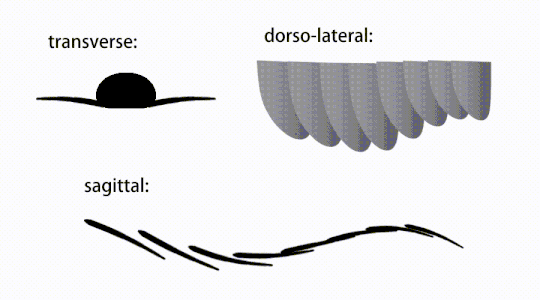
Bevægelsen af radiodont flagrende vedhæng

Ventralt billede af en generaliseret GLS-bærende radiodont, der viser gnathobase-lignende strukturer (GLS'er) forbundet med reducerede forreste flapper



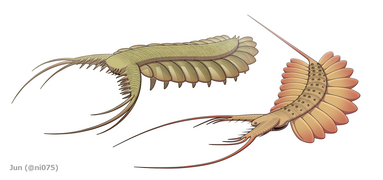
Bagstammevedhængene var finnelignende kropsflapper ('laterale klapper' eller 'lapper' i nogle undersøgelser), sædvanligvis et par ventrale klapper pr. kropssegment, hver lidt overlappende den ene mere fortil, men yderligere, ikke-overlappende sæt af smaa Rygflapper kan forekomme hos nogle Hurdiid- arter. Flapperne kan have adskillige venelignende strukturer (omtalt som 'forstærkende stråler', 'klapstråler', 'tværgående stænger', 'tværgående linjer' eller 'årer'). Flapperne på halsområdet (benævnt 'reducerede flapper', 'nakkeflapper', 'hovedflapper', 'forreste flapper' eller 'differentierede flapper') er betydeligt reduceret. Hos nogle arter opstod kæbelignende fodringsvedhæng kaldet gnathobase-lignende strukturer (GLS'er) fra hver af bunden af deres reducerede nakkeflapper. Talrige aflange bladlignende forlængelser (benævnt lancetformede blade eller lameller) arrangeret i en række og danner bånd af gællelignende strukturer kendt som setalblade, dækkede den dorsale overflade af hvert kropssegment. I det mindste i Aegirocassis er hvert af de lancetformede blade dækket af rynker. Bukflapperne kan være homologe med endopoden af de biramøse lemmer af euarthropoder og lobopodøse lemmer (lobopods) af gællede lobopodier , og de dorsale flaps og setalblade kan være homologe med de udgangs- og gællebærende rygflapper fra den tidligere taxa. Stammen kan ende enten med en halevifte bestående af 1 til 3 par blade, et par lange furcae, en aflang terminal struktur eller en kendeløs stump spids.

Stanleycaris , Hurdia , Aegirocassis , Peytoia og Cambroraster er alle eksempler på hurdiid radiodonts.

Anomalocaris er medlem af familien anomalocarididae , som på et tidspunkt omfattede alle radiodonts, men nu kun omfatter nogle få slægter.

Amplectobelua og Lyrarapax er repræsentanter for amplectobeluidae , som er en meget inkluderende familie af primært kinesiske radiodonter.

"Anomalocaris" briggsi og Tamisiocaris er eksempler på familien tamisiocarididae , som udelukkende var suspensionsføde radiodonter fra Cambrium.



Interne strukturer

Spor af muskler , fordøjelsessystem og nervesystem blev beskrevet fra nogle radiodont-fossiler. Par veludviklede muskler blev forbundet til ventrale klapper placeret ved de laterale hulrum i hvert kropssegment. Mellem de laterale muskler er et sofistikeret fordøjelsessystem, dannet af en udvidelse af for- og bagtarmen, begge forbundet med en smal mellemtarm forbundet med seks par tarmdivercula (fordøjelseskirtler). Sammenlignet med de tre-segmenterede hjerner fra euarthropoder og to-segmenterede hjerner af onychophorans , er hjernen af radiodonts sammensat af kun et hjernesegment, der stammer fra den okulære somit, protocerebrum. Nerverne af de frontale vedhæng og sammensatte øjne opstod fra de forreste og laterale områder af hjernen. Bagved hjernen var et par tilsyneladende usammensmeltede ventrale nervesnore , som løb gennem dyrets halsregion.
Paleoøkologi
Fysiologi

Radiodonts blev fortolket som nektoniske eller nektobentiske dyr, hvor deres morfologi tyder på en aktiv svømmelivsstil. De muskuløse, overlappende bugflapper kan have drevet dyret gennem vandet, muligvis ved at bevæge sig i en bølgelignende formation, der ligner moderne rokker og blæksprutter . Par af rygklapper, som udgør en halevifte hos nogle arter, kan have hjulpet med at styre og/eller stabilisere dyret under bevægelse. I Anomalocaris antyder morfologien af haleviften endda, at den hurtigt kunne ændre sin svømmeretning effektivt. På den anden side har nogle hurdiider funktioner, der er væsentligt specialiserede til en nektobentisk livsstil, såsom Cambroraster med sit kuppellignende H-element, der ligner skjoldet på en hesteskokrabbe . Bånd af setalblade med rynkende lancetformede blade kan have øget overfladearealet, hvilket tyder på, at de var gæller , der sørgede for dyrets åndedrætsfunktion . Overflod af resterne af scleritzede strukturer såsom disartikulerede frontale vedhæng og hovedskleritkomplekser tyder på, at der kan være sket massefyldte hændelser blandt radiodonter , en adfærd, som også er blevet rapporteret hos nogle andre kambriske leddyr såsom trilobitter .
Kost






Radiodonts havde forskellige fodringsstrategier, som kunne kategoriseres som rovdyr , sedimentsigter eller suspension, filterfødere . For eksempel kunne rovdyr fra rovdyr som Anomalocaris og Amplectobeluids have været i stand til at fange adrætte byttedyr ved at bruge deres rovdyrs frontalvedhæng; sidstnævnte bar endda en robust endit til at holde bytte som en tang . Med det mindre hovedskjoldkompleks og store overflade af arthrodiale membraner havde frontale vedhæng af disse taxa større fleksibilitet. Kraftige frontale vedhæng af sedimentsigter som Hurdia og Peytoia har takkede enditter med mesial krumning, som kunne danne en kurvlignende fælde til at rive gennem sediment og føre fødeemner mod den veludviklede orale kegle. Enditter af frontale vedhæng fra suspensions-/filterfødere som Tamisiocaris og Aegirocassis har fleksible, tætpakkede hjælperygsøjler, som kan frafiltrere organiske komponenter såsom mesozooplankton og fytoplankton ned til 0,5 mm. Frontale vedhæng af Caryosyntrips , som er usædvanlige for radiodonts ved at have retningen af enditbærende overflader modsat hinanden og kan have været i stand til at manipulere og knuse bytte i en sakse-lignende udskærings- eller gribebevægelse.
Orale kegler af radiodonts kan have været brugt til sugning og/eller bid. Sammen med den store variation af frontale vedhæng i forskellige arter af radiodonts, antyder differentiering af orale kegler mellem arter også præferencer for forskellige diæter. For eksempel kan den triradiale orale kegle af Anomalocaris med uregelmæssige, tuberkulære tandplader og en lille åbning være blevet tilpasset til små og nektoniske byttedyr, ; mens de stive tetraradiale orale kegler af Peytoia , Titanokorys , Hurdia og en isoleret oral kegle tilskrevet Cambroraster med en større åbning og nogle gange yderligere tandplader kan have været i stand til at indtage større fødeemner i forhold til deres kropsstørrelse og sandsynligvis bentiske eller endobentiske bytte. .
Klassifikation
Taksonomiske affiniteter
|
|||||||||||||||||||||||||||||||||||||||||||||
| Opsummeret fylogeni mellem Radiodonta og andre Ecdysozoan taxa |






Opabiniiderne Opabinia (øverst) og Utaurora ( nederst )
Radiodont/ opabiniid - euarthropod mellemliggende Kylinxia

En diagrammatisk rekonstruktion af den basale deuteropod Erratus


De fleste fylogenetiske analyser tyder på, at radiodonter sammen med opabiniider ( Opabinia og Utaurora ) er stammegruppe leddyr lige basale til deuteropoda , en klade, der inkluderer øvre stilk (fx fuxianhuiids og toskallede leddyr) og krone Euarthropoda (f.eks . Chelicatandi , M , og M ). Denne fortolkning understøttes af adskillige leddyr-grundplaner fundet på radiodonts og opabiniider, såsom stilkede sammensatte øjne , fordøjelseskirtler, trunkvedhæng dannet af dorsale og ventrale elementer (forløber for leddyrs biramøse vedhæng). Sammenlignet med opabiniider, som har posterior mundåbning og sammensmeltede frontal-næsten vedhæng (sammenlignelig med euarthropod posterior-vendende labrum /hypostom-kompleks), havde radiodonter på den anden side euarthropod-lignende dorsal sklerit (H-element) og arthropodization (frontale vedhæng) på deres hovedregioner sammen med kutikulære tarmtermini. Det faktum, at både radiodonter og opabiniider mangler eksoskelet på deres kropsregion, tyder på, at oprindelsen af sammensatte øjne og arthropodisering (segmenterede vedhæng) går forud for arthrodization (fuldt sæt af trunk exoskelet) i leddyrstammen. Den indsnævrede nakkeregion med fodrende appendikulære strukturer af nogle radiodont kan også kaste lys over oprindelsen af det sofistikerede leddyrhoved, som blev dannet ved sammensmeltning af flere forreste kropssegmenter. Basale deuteropoder, der har en blanding af radiodont/opabiniid-karakterer som Kylinxia og Erratus , kan repræsentere mellemformer mellem radiodonts, opabiniider og andre euarthropoder.

Siberiid-lobopodierne Siberion (øverst til venstre), Megadictyon (nederst i midten) og Jianshanopodia (øverst til højre)
De ' gællede lobopodier ' Pambdelurion (venstre) og Kerygmachela (højre)

Megacheira eller 'store appendage arthropoder', en klasse af mulige stamchelicerat , der tidligere blev anset for at være radiodonts nære slægtning



Taxa lige basal til radiodont-, opabiniid- og euarthropod-grenen er ' gællede lobopodier ' som Pambdelurion og Kerygmachela , som lejlighedsvis forenede sig under klassen Dinocaridida med opabibiider og radiodonts. De har kropsflapper, fordøjelseskirtler, store (formodentlig sammensatte) øjne og specialiserede frontale vedhæng som de tidligere taxaer, men deres frontale vedhæng er ikke arthropodized eller fusionerede, øjne siddende, gællelignende strukturer mindre fremtrædende, og har helt sikkert lobopod under hver af deres klapper. Taxa selv basal til 'gællede lobopodier' er siberiider som Megadictyon og Jianshanopodia , en gruppe af lobopodier bar robuste frontale vedhæng og fordøjelseskirtler, men ingen kropsflapper. Sådanne mellemformer mellem lobopodian og radiodont/euarthropod tyder på, at den samlede gruppe Arthropoda opstod fra en parafyletisk lobopodian-grad sammen med de to andre eksisterende panarthropod phyla Tardigrada og Onychophora .
Tidligere undersøgelser kan antyde radiodonter som en anden gruppe end stamme-leddyr, såsom en hidtil ukendt phylum ; cycloneuralia n orme, der er gennemgået konvergent med leddyr (baseret på de cycloneural-lignende radiale munddele); stamme chelicerate euarthropoder sideløbende med megacheira ns også kendt som store appendage arthropoder (baseret på ligheden mellem radiodont frontale appendage, megacheiran store appendages og chelicerae ); eller Schinderhannes bartelsi , som i nyere analyser opløste sig som en hurdiid radiodont, som en art, der er tættere beslægtet med euarthropoder end andre radiodonts (baseret på nogle formodede euarthropod-lignende træk fundet på Schinderhannes ). Men ingen af dem blev understøttet af senere undersøgelser. De radiale munddele er ikke cykloneuralsk-eksklusive og er mere sandsynligt et resultat af konvergent evolution eller ecdysozoa n plesimorphy , da de også er blevet fundet i panarthropoder såsom tardigrad og nogle lobopodier ; radiodonts, der manglede definitive euarthropod-træk såsom trunk tergites og multiple hovedvedhæng, og de megacheiranske store vedhæng blev anset for at være deutocerebrale, som kunne være ikke-homologe med radiodont protocerebrale frontale vedhæng; formodede euarthropod-karakterer fundet på det enkelte Schinderhannes -fossil er tvivlsomt og kan præsentere andre radiodont-lignende strukturer.
Indbyrdes forhold
|
|||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
| Phylogeny of Radiodonta efter Moysiuk & Caron 2021 |





Traditionelt er alle radiodont-arter blevet placeret i én familie, Anomalocarididae , deraf det tidligere almindelige navn 'anomalocaridid', og det blev stadig lejlighedsvis brugt til at henvise til hele rækkefølgen selv efter omklassificering. Siden omplaceringen udført af Vinther et al. 2014 blev de fleste af radiodont-arterne omklassificeret inden for tre nye familier: Amplectobeluidae , Tamisiocarididae (tidligere Cetiocaridae ) og Hurdiidae . Inklusive Anomalocarididae, kan de fire nyere radiodont-familier danne kladen Anomalocarida .
Den originale beskrivelse af ordenen Radiodonta omfattede Anomalocaris , Laggania (senere kendt som Peytoia ), Hurdia , Proboscicaris , Amplectobelua , Cucumericrus og Parapeytoia . Imidlertid betragtes Proboscicaris nu som et juniorsynonym for Hurdia , og Parapeytoia anses for at være en Megacheiran . På grund af den begrænsede opdagelse er positionen af Cucumericrus i Radiodonta uklar, da den enten blev udelekteret ved fylogenetisk analyse eller løst i en polytomi med Radiodonta og Euarthropoda .
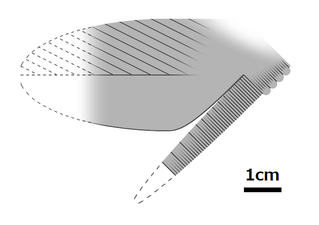
En af de dårligt kendte kropsdele (stammevedhæng) af Cucumericrus decoratus ; denne art repræsenterer muligvis ikke en ægte radiodont.

Frontal vedhæng af "Anomalocaris" briggsi . På trods af det foreslåede generiske navn er det usandsynligt, at denne radiodont er en art af slægten Anomalocaris baseret på fylogenetisk analyse.

Den første dybdegående fylogenetiske analyse af Radiodonta blev udført af Vinther et al . i 2014, efterfulgt af en håndfuld subsequest-studier med mere eller mindre modificerede resultater. I de fleste analyser er Caryosyntrips den basal-mest slægt, men opløst i en polytomi med andre radiodonts og Euarthropoda (sammen med Cucumericrus, hvis inkluderet). Med udelukkelse af Caryosyntrips og Cucumericrus er monofylien af Radiodonta bredt understøttet, med nogle få resultater tyder på mulig paraphyly (enten Anomalocarididae+Amplectobeluidae clade eller Hurdiidae søster til Euarthropoda). Formodede synapomorfier af monofyletisk Radiodonta inklusive tredelt hovedskleritkompleks og differentieret nakkeregion. Slægten Anomalocaris i bredere forstand fandtes altid at være polyfyletisk , sædvanligvis med "Anomalocaris" kunmingensis og "Anomalocaris" briggsi opløst som et medlem af henholdsvis Amplectobeluidae og Tamisiocarididae . Indbyrdes sammenhæng mellem Amplectobeluidae er usikker, da de amplectobeluide affiniteter af Lyrarapax og Ramskoeldia lejlighedsvis blev stillet spørgsmålstegn ved. Monofili af speciose-familien Hurdiidae blev genfundet ved de fleste analyser og godt understøttet af adskillige synapomorfier (f.eks. distalt artikuleret område af frontalt vedhæng med proksimale 5 podomerer, der bærer ulige endititter), med Tamisiocarididae ofte foreslået at være dens søstergruppe.
-
Radiodonta
- ? Cucumericrus (formodet radiodont)
- Caryosyntrips
-
Anomalocarida
- Paranomalocaris (placeret i Anomalocarididae af nogle undersøgelser.)
- Laminacaris (placeret i Amplectobeluidae af nogle undersøgelser.)
- Houcaris (enten placeret i Anomalocarididae, Amplectobeluidae eller Tamisiocarididae.)
- Innovatiocaris
-
Anomalocarididae
- Anomalocaris (i en bredere forstand kan nogle arter placeres i de andre familier.)
- Lenisicaris
-
Amplectobeluidae
- Lyrarapax (position stillet spørgsmålstegn ved af nogle undersøgelser.)
- Amplectobelua
- Ramskoeldia (position sat spørgsmålstegn ved af nogle undersøgelser.)
- "Anomalocaris" kunmingensis (ikke - Anomalocaris - arter venter på at blive omdøbt. Mærket som Amplectobelua kunmingensis af Vinther et al. 2014.)
-
Tamisiocarididae
- Tamisiocaris
- "Anomalocaris" briggsi (ikke - Anomalocaris - arter venter på at blive omdøbt.)
-
Hurdiidae
- Peytoia
- Schinderhannes
- Hurdia
- Aegirocassis
- Stanleycaris
- Pahvantia
- Ursulinacaris
- Cambroraster
- ? Zhenghecaris (formodet radiodont)
- Cordatacaris
- Buccaspinea
- Titanokorys
| Arter | Original beskrivelse | Årstal navngivet | Familie | Alder | Beliggenhed | Frontal vedhæng | Hovedskleritkompleks |
|---|---|---|---|---|---|---|---|
| Cucumericrus decoratus | Hou, Bergström og Ahlberg | 1995 | (ikke tildelt) | Kambrium fase 3 |
|
Ukendt | Ukendt |
| Caryosyntrips serratus | Daley & Budd | 2010 | (ikke tildelt) | Wuliuan – Drumian |
|

|
Ukendt |
| Caryosyntrips camurus | Pates & Daley | 2017 | (ikke tildelt) | Wuliuan |
|

|
Ufuldstændig |
| Caryosyntrips durus | Pates & Daley | 2017 | (ikke tildelt) | Drumian |
|

|
Ukendt |
| Paranomalocaris multisegmentalis | Wang, Huang og Hu | 2013 | Anomalocarididae ? | Kambrium fase 4 |
|

|
Ukendt |
| Paranomalocaris simplex | Jiao, Pates, Lerosey-Aubril, Ortega-Hernandez, Yang, Lan, Zhang | 2021 | Anomalocarididae ? | Kambrium fase 4 |
|

|
Ukendt |
| Laminacaris chimera | Guo, Pates, Cong, Daley, Edgecombe, Chen og Hou | 2018 | (kontroversiel) | Kambrium fase 3 |
|

|
Ukendt |
| Innovatiocaris maotianshanensis | Zeng, Zhao, Zhu | 2022 | (ikke tildelt) | Kambrium fase 3 |
|

|
Ufuldstændig |
| Innovatiocaris ? multispiniformis | Zeng, Zhao, Zhu | 2022 | (ikke tildelt) | Kambrium fase 3 |
|

|
Ukendt |
| Anomalocaris canadensis | Whiteaves | 1892 | Anomalocarididae | Wuliuan |
|

|

|
| Lenisicaris pennsylvanica (tidligere Anomalocaris pennsylvanica ) | Resser | 1929 | Anomalocarididae | Kambrium fase 3 |
|

|
Ukendt |
| Lenisicaris lupata | Wu, Ma, Lin, Sun, Zhang og Fu | 2021 | Anomalocarididae | Kambrium fase 3 |
|

|
Ukendt |
| "Anomalocaris" kunmingensis | Wang, Huang og Hu | 2013 | Amplectobeluidae | Kambrium fase 4 |
|

|
Ukendt |
| Houcaris magnabasis (tidligere Anomalocaris magnabasis ) | Pates, Daley, Edgecombe, Cong & Lieberman | 2019 | (kontroversiel) | Kambrium fase 4 |
|

|
Ukendt |
| Houcaris saron (tidligere Anomalocaris saron ) | Hou, Bergström og Ahlberg | 1995 | (kontroversiel) | Kambrium fase 3 |
|

|
Ukendt |
| "Anomalocaris" briggsi | Nedin | 1995 | Tamisiocarididae | Kambrium fase 4 |
|

|
Ukendt |
| Ramskoeldia platyacantha | Cong, Edgecombe, Daley, Guo, Pates og Hou | 2018 | Amplectobeluidae | Kambrium fase 3 |
|

|
Ufuldstændig |
| Ramskoeldia consimilis | Cong, Edgecombe, Daley, Guo, Pates og Hou | 2018 | Amplectobeluidae | Kambrium fase 3 |
|

|
Ufuldstændig |
| Lyrarapax unguispinus | Cong, Ma, Hou, Edgecombe og Strausfield | 2014 | Amplectobeluidae | Kambrium fase 3 |
|

|
|
| Lyrarapax trilobus | Cong, Daley, Edgecombe, Hou og Chen | 2016 | Amplectobeluidae | Kambrium fase 3 |
|

|
|
| Amplectobelua symbrachiata | Hou, Bergström og Ahlberg | 1995 | Amplectobeluidae | Kambrium fase 3 |
|

|

|
| Amplectobelua stephenensis | Daley & Budd | 2010 | Amplectobeluidae | Wuliuan |
|

|
Ukendt |
| Tamisiocaris borealis | Daley & Peel | 2010 | Tamisiocarididae | Kambrium fase 3 |
|

|
Ufuldstændig |
| Ursulinacaris grallae | Pates, Daley & Butterfield | 2019 | Hurdiidae | Wuliuan |
|

|
Ukendt |
| Schinderhannes bartelsi | Kühl, Briggs og Rust | 2009 | Hurdiidae | Emsian |
|
Ufuldstændig | Ufuldstændig |
| Stanleycaris hirpex | Pates, Daley og Ortega-Hernández | 2018 | Hurdiidae | Wuliuan |
|

|
P-element er ukendt, muligvis fraværende |
| Peytoia nathorsti | Walcott | 1911 | Hurdiidae | Wuliuan – Drumian |
|

|
Ufuldstændig |
| Peytoia infercambriensis (tidligere Cassubia infercambriensis ) | Lendzion | 1975 | Hurdiidae | Kambrium fase 3 |
|

|
Ukendt |
| Aegirocassis benmoulai | Van Roy, Daley og Briggs | 2015 | Hurdiidae | Tremadocian |
|

|

|
| Hurdia victoria | Walcott | 1912 | Hurdiidae | Wuliuan – Drumian |
|

|

|
| Hurdia triangulata | Walcott | 1912 | Hurdiidae | Wuliuan |
|
|

|
| Cambroraster falcatus | Moysiuk og Caron | 2019 | Hurdiidae | Wuliuan |
|

|

|
| Pahvantia hastata | Robison og Richards | 1981 | Hurdiidae | Drumian |
|

|

|
| Cordaticaris striatus | Sun, Zeng og Zhao | 2020 | Hurdiidae | Drumian |
|
Ufuldstændig |

|
| Zhenghecaris shankouensis | Vanner, Chen, Huang, Charbonnier og Wang | 2006 | Hurdiidae | Kambrium fase 3 |
|
Ukendt |
 |
| Buccaspinea cooperi | Pates, Lerosey-Aubril, Daley, Kier, Bonino & Ortega-Hernández | 2021 | Hurdiidae | Drumian |
|

|
Ukendt |
| Titanokorys gainesi | Caron og Moysiuk | 2021 | Hurdiidae | Wuliuan |
|

|

|
Historie


Frontal vedhæng af Anomalocaris canadensis

Oral kegle af Peytoia nathorsti

H-element af Hurdia victoria
![Parrede frontale vedhæng fra en unavngiven hurdiid radiodont[36]](https://upload.wikimedia.org/wikipedia/commons/thumb/1/1d/USNM_PAL_57490.jpg/267px-USNM_PAL_57490.jpg)
Parrede frontale vedhæng fra en unavngiven hurdiid radiodont



Historien om radiodonts er kompleks. Ufuldstændige prøver vedrørende forskellige kropsdele af den samme art var historisk blevet fortolket som tilhørende forskellige arter og endda forskellige phyla. Forud for deres anerkendelse som en gruppe, var radiodont-prøver blevet tildelt fem forskellige phyla: Porifera , Cnidaria , Echinodermata , Annelida og Arthropoda .
De første kendte radiodont-prøver blev indsamlet fra trilobite-sengene på Mount Stephen af Richard G. McConnell fra Geological Survey of Canada i 1886 eller 1888. Disse prøver blev navngivet Anomalocaris canadensis i 1892 af GSC-palæontolog Joseph Whiteaves . Whiteaves fortolkede prøverne, der nu er kendt for at være isolerede frontale vedhæng, som underlivet på et phyllocarid krebsdyr . Yderligere radiodont-prøver blev beskrevet i 1911 af Charles Walcott . Han fortolkede en isoleret oral kegle, som han kaldte Peytoia nathorsti , som en vandmand, og et dårligt bevaret, men relativt komplet eksemplar, som han kaldte Laggania cambria , som en holothurian. I 1912 navngav Walcott Hurdia victoria og H. triangulata baseret på isolerede H-elementer, som han fortolkede som skjoldene af krebsdyr. Isolerede frontale vedhæng af Peytoia og Hurdia , samlet kendt som "Appendage F" i Briggs 1979, blev alle identificeret som dem fra Sidneyia på det tidspunkt. Et Hurdia P-element blev navngivet Proboscicaris i 1962 og fortolket som skjoldet af en toskallet leddyr.
The Geological Survey of Canada indledte en revision af Burgess Shale fossiler i 1966, overvåget af Cambridge University palæontolog Harry B. Whittington . Denne revision ville i sidste ende føre til opdagelsen af den komplette radiodont kropsplan. I 1978 anerkendte Simon Conway Morris , at Lagganias munddele var Peytoia- lignende, men han tolkede dette som bevis på, at det var et sammensat fossil bestående af en Peytoia- vandmand og en svamp. I 1979 erkendte Derek Briggs , at fossilerne af Anomalocaris var vedhæng, ikke underliv, men fortolkede dem som gåben ved siden af "vedhæng F". Det var først i 1985, at den sande natur af fossilerne af Anomalocaris , Laggania og Peytoia blev anerkendt, og de blev alle tildelt en enkelt slægt, Anomalocaris . Efterfølgende blev det erkendt, at Anomalocaris var en adskilt form fra de to andre, hvilket resulterede i en opdeling i to slægter, hvoraf sidstnævnte på forskellige måder blev navngivet Laggania og Peytoia, indtil det blev fastslået, at begge repræsenterede den samme art, og Peytoia havde prioritet. Det blev senere erkendt, at nogle af de fossiler, der blev tildelt disse taxaer, tilhørte en anden form, som blev anerkendt som bærende et skjold bestående af Hurdia- og Proboscicaris - elementer. Endelig, i 2009, blev disse eksemplarer omskrevet som Hurdia . Selv efter disse genkendelser var delvise fejlidentifikationer (f.eks. oral kegle og frontale vedhæng af Peytoia blevet tildelt til henholdsvis Anomalocaris og Hurdia ) også blevet afsløret af efterfølgende undersøgelser.
Selve taxonet Radiodonta blev opfundet i 1996 af Desmond Collins, efter at det blev fastslået, at Anomalocaris og dets slægtninge repræsenterede en karakteristisk slægt med leddyr-tilhørsforhold snarere end en hidtil ukendt filum. Collins etablerede også klassen Dinocarida til at indeholde ordenen Radiodonta såvel som Opabiniidae , som han anerkendte som distinkt på grund af dens manglende karakteristiske orale keglestruktur af radiodonts. Radiodonta fik først en fylogenetisk definition i 2014. Radiodonta blev oprindeligt anset for at indeholde en enkelt familie, Anomalocarididae , men den blev opdelt i fire familier i 2014: Amplectobeluidae , Anomalocarididae, Cetiocaridae og Hurdiidae . Navnet Cetiocaridae var ikke i overensstemmelse med den internationale kodeks for zoologisk nomenklatur og blev derfor omdøbt til Tamisiocarididae i 2019.
Indtil 2010'erne blev radiodonter typisk anset for at være ensartet store apex-rovdyr, men opdagelser af nye arter i løbet af det årti førte til en betydelig stigning i den kendte økologiske og morfologiske mangfoldighed af gruppen.