
Pericyte - Pericyte
| Pericyte | |
|---|---|

Transmission elektronmikrografi af en mikrokar viser pericytter, der beklæder den ydre overflade af endotelceller , der omkranser en erythrocyt (E).
| |
| detaljer | |
| Identifikatorer | |
| Latin | pericytus |
| MeSH | D020286 |
| TH | H3.09.02.0.02006 |
| FMA | 63174 |
|
Anatomiske termer for mikroanatomi | |
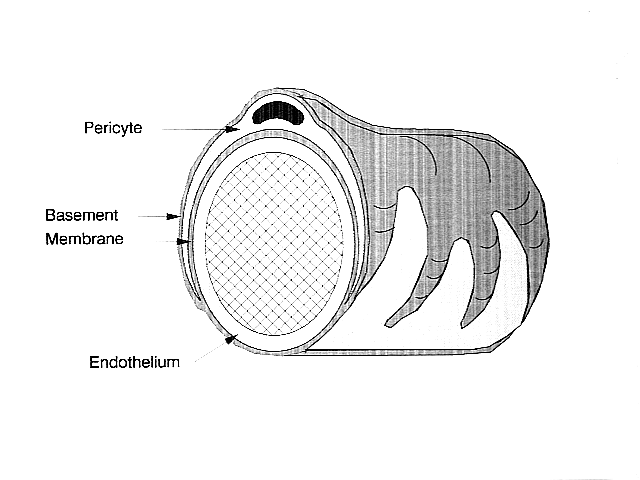
Pericytter (tidligere kendt som Rouget-celler ) er multifunktionelle vægmalerceller i mikrocirkulationen, der vikles rundt om endotelcellerne, der fører kapillærerne gennem hele kroppen. Pericytter er indlejret i basalmembranen i blodkapillærer, hvor de kommunikerer med endotelceller ved hjælp af både direkte fysisk kontakt og parakrin signalering . Moricologien, fordelingen, densiteten og molekylære fingeraftryk af pericytter varierer mellem organer og vaskulære senge. Pericytter hjælper med at opretholde homøostatiske og hæmostatiske funktioner i hjernen , et af organerne med højere pericytdækning, og opretholder også blod -hjerne -barrieren . Disse celler er også en nøglekomponent i den neurovaskulære enhed , som omfatter endotelceller, astrocytter og neuroner . Pericytter er blevet postuleret for at regulere kapillær blodgennemstrømning og clearance og fagocytose af cellulært affald in vitro. Pericytter stabiliserer og overvåger modning af endotelceller ved hjælp af direkte kommunikation mellem cellemembranen såvel som via parakrin signalering. Mangel på pericytter i centralnervesystemet kan forårsage øget permeabilitet af blod -hjerne -barrieren.
Struktur

I centralnervesystemet (CNS) vikles pericytter rundt om endotelcellerne, der beklæder kapillærens inderside. Disse to celletyper kan let skelnes fra hinanden baseret på tilstedeværelsen af pericytens fremtrædende runde kerne sammenlignet med endotelcellernes flade aflange kerne. Pericytter projekterer også fingerlignende forlængelser, der vikler rundt om kapillærvæggen, så cellerne kan regulere kapillær blodgennemstrømning.
Både pericytter og endotelceller deler en basalmembran, hvor der laves en række intercellulære forbindelser. Mange typer integrinmolekyler letter kommunikationen mellem pericytter og endotelceller adskilt af basalmembranen. Pericytter kan også danne direkte forbindelser til naboceller ved at danne pinde- og sokkelarrangementer, hvor dele af cellerne hænger sammen, svarende til tandhjulene på et ur. Ved disse indgribende sites, gap junctions kan dannes, som tillader pericytterne og naboceller til at udveksle ioner og andre små molekyler. Vigtige molekyler i disse intercellulære forbindelser omfatter N-cadherin , fibronectin , connexin og forskellige integriner.
I nogle områder af basalmembranen kan der findes adhæsionsplakker sammensat af fibronectin. Disse plaketter letter forbindelsen af basalmembranen til den cytoskeletale struktur sammensat af actin og plasmamembranen i pericytterne og endotelcellerne.
Fungere
Skeletmuskulaturgenerering og fedtdannelse
Pericytter i den skeletstrimlede muskel er af to forskellige populationer, hver med sin egen rolle. Den første pericyt subtype (Type-1) kan differentiere sig til fedtceller , mens den anden (Type-2) til muskelceller. Type-1 karakteriseret ved negativ ekspression for nestin (PDGFRβ+CD146+Nes-) og type-2 karakteriseret ved positiv ekspression for nestin (PDGFRβ+CD146+Nes+). Mens begge typer er i stand til at formere sig som reaktion på glycerol eller BaCl2-induceret skade, giver type-1 pericytter kun adipogene celler som reaktion på glycerolinjektion, og type-2 bliver myogen som reaktion på begge typer skader . I hvilket omfang type-1 pericytter deltager i fedtophobning vides ikke.
Angiogenese og overlevelse af endotelceller
Pericytter er også forbundet med differentiering og multiplikation af endotelceller, angiogenese , overlevelse af apoptotiske signaler og rejser. Visse pericytter, kendt som mikrovaskulære pericytter, udvikler sig omkring kapillærernes vægge og hjælper med at tjene denne funktion. Mikrovaskulære pericytter er muligvis ikke kontraktile celler, da de mangler alfa- aktin isoformer, strukturer, der er almindelige blandt andre kontraktile celler. Disse celler kommunikerer med endotelceller via gap junctions og får igen endotelceller til at formere sig eller blive selektivt hæmmet. Hvis denne proces ikke forekom, kan der opstå hyperplasi og unormal vaskulær morfogenese . Disse typer af pericyter kan også fagocytose eksogene proteiner. Dette tyder på, at celletypen kan have været afledt af mikroglia .
Et slægtforhold til andre celletyper er blevet foreslået, herunder glatte muskelceller , neurale celler, NG2 glia , muskelfibre , adipocytter samt fibroblaster og andre mesenkymale stamceller . Hvorvidt disse celler differentierer til hinanden er imidlertid et fremragende spørgsmål i feltet. Pericytternes regenerative kapacitet påvirkes af aldring. Sådan alsidighed er nyttig, da de aktivt ombygger blodkar i hele kroppen og derved kan blande homogent med det lokale vævsmiljø .
Bortset fra at skabe og ombygge blodkar, har pericytter vist sig at beskytte endotelceller mod døden via apoptose eller cytotoksiske elementer. Det er blevet vist in vivo, at pericytter frigiver et hormon kendt som pericytisk aminopeptidase N/pAPN, der kan bidrage til at fremme angiogenese. Da dette hormon blev blandet med cerebrale endotelceller såvel som astrocytter, grupperede pericytterne sig i strukturer, der lignede kapillærer. Når den eksperimentelle gruppe endvidere indeholdt alt det følgende med undtagelse af pericytter, ville endotelcellerne undergå apoptose. Det blev således konkluderet, at pericytter skal være til stede for at sikre, at endotelceller fungerer korrekt, og astrocytter skal være til stede for at sikre, at begge forbliver i kontakt. Hvis ikke, kan der ikke forekomme korrekt angiogenese. Det har også vist sig, at pericytter bidrager til overlevelse af endotelceller, da de udskiller proteinet Bcl-w under cellulær krydstale. Bcl-w er et instrumentalt protein i vejen, der håndhæver VEGF-A- ekspression og modvirker apoptose. Selvom der er nogle spekulationer om, hvorfor VEGF er direkte ansvarlig for at forhindre apoptose, menes det at være ansvarlig for modulering af apoptotiske signaltransduktionsveje og hæmning af aktivering af apoptose-inducerende enzymer . To biokemiske mekanismer anvendt af VEGF til at opnå dette ville være phosphorylering af ekstracellulær regulatorisk kinase 1 (ERK-1, også kendt som MAPK3), som opretholder celleoverlevelse over tid og inhibering af stressaktiveret proteinkinase/c-jun-NH2 kinase , som også fremmer apoptose.
Blod hjerne barrieren
Pericytter spiller en afgørende rolle i dannelsen og funktionaliteten af blod -hjerne -barrieren . Denne barriere består af endotelceller og sikrer beskyttelse og funktionalitet af hjernen og centralnervesystemet. Det har vist sig, at pericytter er afgørende for postnatal dannelse af denne barriere. Pericytter er ansvarlige for dannelse af tæt kryds og handel med vesikler blandt endotelceller. Desuden tillader de dannelsen af blod -hjerne -barrieren ved at hæmme virkningerne af CNS -immunceller (som kan skade dannelsen af barrieren) og ved at reducere ekspressionen af molekyler, der øger vaskulær permeabilitet.
Bortset fra dannelse af blod -hjerne -barriere spiller pericytter også en aktiv rolle i dets funktionalitet. Dyremodeller for udviklingstab af pericytter viser øget endotel-transcytose samt skæv arterio-venøs zonering, øget ekspression af leukocytadhæsionsmolekyler og mikroaneurismer. Tab eller dysfunktion af pericytter er også teoretiseret til at bidrage til neurodegenerative sygdomme som Alzheimers , Parkinsons og ALS gennem nedbrydning af blod-hjerne-barrieren.
Blodgennemstrømning
Stigende bevis tyder på, at pericytter kan regulere blodgennemstrømningen på kapillærniveau. For nethinden er der blevet offentliggjort film, der viser, at pericytter indsnævrer kapillærer, når deres membranpotentiale ændres for at forårsage calciumtilstrømning, og i hjernen er det blevet rapporteret, at neuronal aktivitet øger lokal blodgennemstrømning ved at få pericytter til at udvide kapillærer, før opstrøms arterioldilatation forekommer . Dette område er kontroversielt, med en nylig undersøgelse, der hævder, at pericytter ikke udtrykker kontraktile proteiner og ikke er i stand til at sammentrække in vivo, selvom sidstnævnte papir er blevet kritiseret for at bruge en meget ukonventionel definition af pericyte, som eksplicit udelukker kontraktile pericytter. Det ser ud til, at forskellige signalveje regulerer indsnævring af kapillærer med pericytter og af arterioler af glatte muskelceller.
Pericytter er vigtige for at opretholde cirkulationen. I en undersøgelse, der omfattede voksne mus med mangel på pericyte, blev cerebral blodgennemstrømning reduceret med samtidig vaskulær regression på grund af tab af både endothelia og pericytter. Signifikant større hypoksi blev rapporteret i hippocampus hos mus med mangel på pericyte samt betændelse og nedsat indlæring og hukommelse .
Klinisk betydning
På grund af deres afgørende rolle i opretholdelse og regulering af endotelcellestruktur og blodgennemstrømning ses abnormiteter i pericytefunktion i mange patologier. De kan enten være til stede i overskud, hvilket fører til sygdomme såsom hypertension og tumordannelse eller i mangel, hvilket fører til neurodegenerative sygdomme.
Hemangiom
De kliniske faser af hæmangiom har fysiologiske forskelle, korreleret med immunofenotypiske profiler af Takahashi et al. I den tidlige proliferative fase (0–12 måneder) udtrykker tumorerne prolifererende cellekerne -antigen (pericytesna), vaskulær endotelvækstfaktor (VEGF) og kollagenase type IV, de to tidligere lokaliseret til både endotel og pericytter, og den sidste til endotel . De vaskulære markører CD31, von Willebrand -faktor (vWF) og glatmuskelaktin (pericytmarkør) er til stede under de prolifererende og involverende faser, men går tabt, efter at læsionen er fuldt involveret.
Hæmangiopericytom

Hemangiopericytoma er en sjælden vaskulær neoplasma eller unormal vækst, der enten kan være godartet eller ondartet. I sin ondartede form kan der forekomme metastaser i lungerne, leveren, hjernen og ekstremiteterne. Det manifesterer sig oftest i lårbenet og det proximale skinneben som et knoglesarkom og findes normalt hos ældre individer, selvom der er fundet tilfælde hos børn. Hemangiopericytoma er forårsaget af overdreven lagdeling af ark med pericytter omkring forkert dannede blodkar. Diagnose af denne tumor er vanskelig på grund af manglende evne til at skelne pericytter fra andre celletyper ved hjælp af lysmikroskopi. Behandlingen kan omfatte kirurgisk fjernelse og strålebehandling, afhængigt af knogletrængningsniveauet og stadiet i tumorens udvikling.
Diabetisk retinopati
Retina hos diabetiske individer udviser ofte tab af pericytter, og dette tab er en karakteristisk faktor for de tidlige stadier af diabetisk retinopati . Undersøgelser har fundet ud af, at pericytter er afgørende for diabetikere for at beskytte endotelcellerne i retinale kapillærer. Med tabet af pericytter dannes mikroaneurismer i kapillærerne. Som svar øger nethinden enten sin vaskulære permeabilitet, hvilket fører til hævelse af øjet gennem et makulaødem eller danner nye kar, der trænger ind i øjets glaslegememembran . Slutresultatet er nedsat eller tab af syn. Selvom det er uklart, hvorfor pericytter går tabt hos diabetespatienter, er en hypotese, at toksisk sorbitol og avancerede glycation-slutprodukter (AGE) ophobes i pericytterne. På grund af opbygningen af glucose, den polyol pathway øger sin flux, og intracellulær sorbitol og fruktose akkumulere. Dette fører til osmotisk ubalance, hvilket resulterer i celleskader. Tilstedeværelsen af høje glukoseniveauer fører også til opbygning af AGE'er, som også skader celler.
Neurodegenerative sygdomme
Undersøgelser har fundet ud af, at pericyttab i den voksne og aldrende hjerne fører til afbrydelse af korrekt cerebral perfusion og vedligeholdelse af blod -hjerne -barrieren, hvilket forårsager neurodegeneration og neuroinflammation. Apoptosen af pericytter i den aldrende hjerne kan være et resultat af en fejl i kommunikationen mellem vækstfaktorer og receptorer på pericytter. Trombocyt-afledt vækstfaktor B ( PDGFB ) frigives fra endotelceller i hjernens vaskulatur og binder til receptoren PDGFRB på pericytter, hvilket initierer deres proliferation og investering i vaskulaturen.
Immunhistokemiske undersøgelser af humant væv fra Alzheimers sygdom og amyotrofisk lateral sklerose viser tab af pericyter og nedbrydning af blod-hjerne-barrieren. Pericyte-mangelfulde musemodeller (som mangler gener, der koder for trin i PDGFB: PDGFRB-signalkaskade) og har en Alzheimers-forårsagende mutation, har forværret Alzheimers-lignende patologi sammenlignet med mus med normal pericyte-dækning og en Alzheimers-forårsagende mutation.
Slag
I tilfælde af slagtilfælde indsnævrer pericytter hjernekapillærer og dør derefter, hvilket kan føre til et langvarigt fald i blodgennemstrømningen og tab af blod-hjerne-barrierefunktion, hvilket øger nervecelles død.
Forskning
Endotel- og pericyte -interaktioner
Endotelceller og pericytter er indbyrdes afhængige, og fejl i korrekt kommunikation mellem de to celletyper kan føre til talrige menneskelige patologier.
Der er flere kommunikationsveje mellem endotelcellerne og pericytterne. Den første transformerer vækstfaktor (TGF) signalering, som medieres af endotelceller. Dette er vigtigt for pericytdifferentiering. Angiopoietin 1 og Tie-2 signalering er afgørende for modning og stabilisering af endotelceller. Trombocyt-afledt vækstfaktor (PDGF) pathway-signalering fra endotelceller rekrutterer pericytter, så pericytter kan migrere til udviklende blodkar. Hvis denne vej er blokeret, fører det til pericyte -mangel. Sphingosin-1-phosphat (S1P) signalering hjælper også med rekruttering af pericyter ved kommunikation gennem G-proteinkoblede receptorer . S1P sender signaler gennem GTPaser, der fremmer N-cadherinhandel til endotelmembraner. Denne handel styrker endotelkontakter med pericytter.
Kommunikation mellem endotelceller og pericytter er afgørende. Hæmning af PDGF -vejen fører til pericyte -mangel. Dette forårsager endotelhyperplasi, unormale kryds og diabetisk retinopati. Mangel på pericytter forårsager også en opregulering af vaskulær endotelvækstfaktor (VEGF), hvilket fører til vaskulær lækage og blødning . Angiopoietin 2 kan fungere som en antagonist til Tie-2 og destabilisere endotelcellerne, hvilket resulterer i mindre endotelceller og pericytinteraktion. Dette fører lejlighedsvis til dannelse af tumorer. Ligesom hæmning af PDGF -vejen reducerer angiopoietin 2 niveauer af pericytter, hvilket fører til diabetisk retinopati.
Ardannelse
Normalt er astrocytter forbundet med ardannelsesprocessen i centralnervesystemet og danner glial ar . Det er blevet foreslået, at en undertype pericytter deltager i denne ardannelse på en glialuafhængig måde. Gennem slægtssporingsundersøgelser blev disse undertype pericytter fulgt efter slagtilfælde, hvilket afslørede, at de bidrager til glialarret ved at differentiere sig til myofibroblaster og deponere ekstracellulær matrix. Dette er imidlertid stadig kontroversielt, da nyere undersøgelser tyder på, at celletypen, der følges i disse arundersøgelser, sandsynligvis ikke er pericytter, men fibroblaster.
Bidrag til voksen neurogenese
De nye beviser (fra 2019) tyder på, at neurale mikrovaskulære pericytter, under instruktion fra residente glialceller, omprogrammeres til interneuroner og beriger lokale neuronale mikrokredsløb. Dette svar forstærkes ved samtidig angiogenese.
Se også
- Hæmangiopericytom
- Mesoangioblast
- Diabetisk retinopati forårsaget af død af pericytter
- Liste over humane celletyper, der stammer fra kimlagene
Referencer
eksterne links
- www.stemcellsfreak.com - Pericytter kan bruges til muskelregenerering
- Diagram på udel.edu
{kind=link}